Download
1 / 40
430 likes | 651 Views
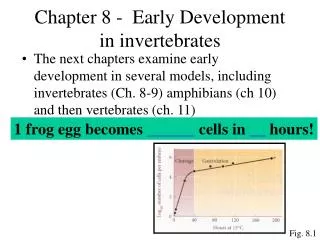
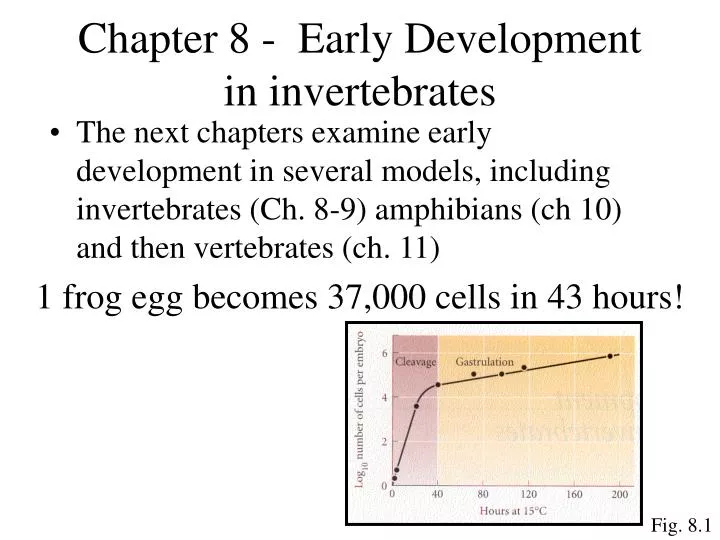
Chapter 8 - Early Development in invertebrates. The next chapters examine early development in several models, including invertebrates (Ch. 8-9) amphibians (ch 10) and then vertebrates (ch. 11). 1 frog egg becomes 37,000 cells in 43 hours!. Fig. 8.1. General Animal Development.

E N D
Chapter 8 - Early Development in invertebrates • The next chapters examine early development in several models, including invertebrates (Ch. 8-9) amphibians (ch 10) and then vertebrates (ch. 11) 1 frog egg becomes 37,000 cells in 43 hours! Fig. 8.1
General Animal Development 2. Gastrulation- Extensive cell rearrangement to form endo-, ecto- and meso-derm • Cleavage- One cell is subdivided into many cells to form a blastula 4. Gameteogenesis- produce germ cells (sperm/egg) Note: Somatic cells denote all non-germ cells 3. Organogenesis- Cells rearranged to produce organs and tissue Recall from lecture 1
How does egg undergo cleavage without increasing it’s size? • Answer- it abolishes G1 and G2 phases of cell cycle • Do you need a cell cycle primer?? • Four cell cycle phases • M- mitosis • G1- Gap 1 • S- DNA Synthesis • G2- Gap 2
Reminder- mitosis occurs in M phase, DNA replication in interphase From Mol. Biol of the Cell by Alberts et al, p864
Cyclin dependent kinases (cdks, cdcs) drive the cell cycle • Cyclins (e.g cycli A, B…) regulate cdk (cdc) activity MPF Example- Mitosis Promoting Factor (MPF= cyclin B+cdc2)
How does egg undergo cleavage without increasing it’s size? • Answer- it abolishes G1 and G2 phases of cell cycle M phase Cyclins are synthesized in eggs Cyclin is degraded S phase
What actually drives the cleavage process? • Answer- Two processes- • 1. Karyokinesis (mitotic division of the nucleus) • The mitotic spindle (microtubules composed of tubulin) does this • 2. Cytokinesis (mitotic division of the cell) • Contractile rings “pinch off” (microfilaments composed of actin) Fig. 8.3 Cytochalasin B prevents cytokineses
General Animal Development 2. Gastrulation- Extensive cell rearrangement to form endo-, ecto- and meso-derm • Cleavage- One cell is subdivided into many cells to form a blastula 4. Gameteogenesis- produce germ cells (sperm/egg) Note: Somatic cells denote all non-germ cells 3. Organogenesis- Cells rearranged to produce organs and tissue Recall from lecture 1
1 2 3 Gastrulation- cells of blastula are dramatically rearranged • Three germ layers are produced Five types of movements
4 5 Gastrulation Five types of movements
Axis formation • Three axes must be determined- • Anterior-posterior (front-back) • Dorsal-ventral (back-belly) • Right-left (right side-left side) Fig. 8.7
Now let’s take a look at one beast- the sea urchin 1. Cleavage • Cleavages 1 and 2 are through animal/vegetal poles 1 2 • Cleavage 4 results in four macromeres and and four micromeres only in vegetal cells 4 3 • Cleavage 3 results in four vegetal and four animal cells Fig. 8.8
Fig. 8.9 • Post cleavage 5 Cell fate map
Micromeres signal other cells via b-catenin to influence fate • Micromere cell fate is autonomous- these become skeletal tissue if placed in a dish • All other cells have conditional specification Example- Transplant micromeres to animal pole at 16 cell stage • Micromeres cause a second invagination • Animal pole cells become vegetal cells Fig. 8.13
Sea urchin (continued) 2. Gastrulation Gastrula Late blastula Egg Later stages Note-micromemeres produce primary mesenchyme which will become larval skeleton Fig. 8.16 Sea urchin development
How do mesenchyme cells know to migrate inside blastocoel? Answer- changing cell attachment proteins Blastocoel Fig. 8.19 Extracellular matrix Basal lamina Hyaline layer 98% decrease in hyaline affinity 100-fold increase in EM/basal lamina affinity Primary mesenchyme cell-
How does invagination occur? Terms- Invagination region is called archenteron Opening created is called the blastopore Answer- swelling of inner lamina layer Vegetal cells secrete chondroitin sulfate proteoglycan, causing inner layer to swell and cause buckling Hyaline Inner layer Outer layer
Now let’s take a look at another creature- the nematode C. elegans • 959 cells at maturity • 1mm long • Produces eggs and sperm (hermaphrodyte) • Transparent • 16 hour from egg to hatch • Entire genome sequences- 19,000 genes What a great model! 1mm
C. elegans 1. Cleavage Oviduct Germ cells undergo mitosis, then begin meiosis as travel down oviduct Cleavage Mature eggs passes through the sperm on the way to the vulva
C. elegans 1. Cleavage • Cleavages 1 produces founder cell (AB) and stem cell (P1) 3 1 2 Fig. 8.42 • Remaining cleavages result in a single stem cell and more founder cells • Cleavage 2 results in three founder cells and one stem cell (P2)
C. elegans How is the anterior-posterior axis determined? • Answer- P-granules- ribonucleoprotein complexes • P-granules always stay associated with the “P” cell 1 3 What directs the P granules? • PAR proteins- these specify polarity, cell division and cytoplasmic localization 5 Fig. 8.43
C. elegans 1. Cleavage • P1 develops autonomously • AB does not (thus is conditional) • What drives P1 lineage? • P granules? No, these do not enter nucleus! • Other possibilities • SKN-1- a bZIP family transcription factor that control EMS cell fate • PAL-1 - a transcription factor required for P2 lineage • PIE-1- inhibits SKN-1 and PAL-1 in P2
Does P2 dictate fate of adjacent cells? Yes- P2 produces a signal that tells ABp to only neurons and hypodermal cells not neurons or pharynx like ABa • GLP-1 is the receptor on ABp, and APX-1 is the ligand on P2 GLP-1 is a Notch family protein APX-1 is a Delta family protein
Chapter 9 - Axis specification in Drosophila • Drosophila genetics is the groundwork for developmental genetics • Cheap, easy to breed and maintain • Drosophila geneticists take pride in being different and in sharing information • Problems- fairly complex, non-transparent Fig. 8.1
1. Cleavage Drosophila • Insects tend to undergo superficial cleavage- cleavage occurs at rim of the egg • In contrast to other creatures, insects form nuclei, then create cells Termed a syncytial blastoderm • Mitotic divisions • #1-#9 - duplicate nuclei (8 min/division 1 7 • Mitotic division • #10 – nuclei migrate to rim • Mitotic division • #11-14 – progressively slower divisions 10 Fig. 9.1
1. Cleavage Drosophila 14 Note – each nuclei has a territory of cytoskeletal proteins • Mitotic divisions • #14 – cells created with each nuclei to create the cellular blastoderm Nuclei staun Tubulin stain Egg plasma membrane folds between nuclei to create individual cells Cycle 11-14- midblastula transition- nuclear division slows and transcription increases Fig. 9.3
2. Gastrulation Ventral Dorsal Ventral furrow (from mesoderm) Fig. 9.6 Segments Head 3 thorax 8 abdominal Fig. 9.7 It becomes the ventral tube
2. Gastrulation Establishment of anterior-posterior polarity-protein gradients rule the day Gene family Examples Maternal effect- in specific region of egg bicoid Nanos caudel Gap- among 1st gene transcribed in embryo Kruppel hunchback Pair rule – result in 7 bands fushi tarazu hairy Segment polarity – result in 14 segments engrailed wingless Fig. 9.8
2. Gastrulation Maternal effect genes Active during creation of syncytial blastoderm nanos mRNA injected, localize to posterior Bicoid mRNA injected in anterior Caudel (diffuse) Hunchback (diffuse) Fig. 9.10 Bicoid prevents caudel mRNA translation Nanos prevents hunchback mRNA translation
Maternal effect genes Oocyte mRNAs Mechanism Anterior Syncytial Blastoderm proteins Posterior Fig. 9.11
Maternal effect genes What if we mess up the bicoid gradient? Inject bicoid into: Wild-type Bicoid- mutant Bicoid-/- Bicoid-/- Wild-type Anterior Middle Posterior Head in middle Two heads Normal Thus, bicoid specifies head development Two tails Fig. 9.14
How does nanos specify posterior? Answer- By preventing hunchback translation Mechanism In anterior, Pumilio binds 3’UTR (untranslated region) of hunchback mRNA, and mRNA is polyadenylated and translated Anterior (no nanos) Posterior (with nanos) In posterior, nanos prevents polyadenyltation, and thus prevents translation Fig. 9.16
2. Gastrulation Gap Pair-rule Segment polarity Determination genes Segmentation genes Egg Specification Determination Bicoid, nanos, hunchback, caudel, etc. Gap genes Pair-rule genes Segment polarity genes Segmentation genes Two steps in Drosophila development (Cell fate is flexible) (Cell fate is determined) Maternal effect genes activate gaps genes, which activate pair-rule genes, which activate segment polarity genes Fig. 9.19 Segmentation genes establish boundaries
Gap a. Gap Genes • Gap genes respond to maternal effect proteins • Gap proteins interact to define specific regions of embryo • Four major gap proteins- hunchback, giant, Kruppel and knirp • These are all DNA binding proteins- activate or repress transcription b. Pair-rule genes • Gap genes activate and repress pair-rule genes in every other stripe, resulting in seven stripes • Three major pair rule proteins- hairy, evenskipped, runt • These are all DNA binding proteins- activate or repress transcription • Cells in each parasegment contains a unique set of pair rule genes expression unlike any other parasegment Fig. 9.21 Pair-rule
b. Pair-rule genes Why do we observe expression of pair-rule proteins in every other segment? Answer- pair-rule genes use different enhancer elements Pair-rule Example- even-skipped (a pair-rule gene) has several enhancers, but only one is active in a given stripe Different concentrations of gap proteins determine pair-rule gene expression This enhancer is only active in stripes #1 Fig. 9.22
c. Segment polarity genes Maternal, gap and pair-rule genes operate before cells are formed Segment polarity Pair-rule syncytial blastoderm 14 Segment polarity genes act once cells are formed Fig. 9.1
c. Segment polarity genes Segment polarity genes encode proteins that make up Hedgehog and Wingless signal transduction pathways One cell produces wingless The adjacent cell produces hedgehog Fig. 9.25 Wingless and hedgehog activate expression of each other
2. Gastrulation Homeotic selector genes Responsible for directing structure formation of each segment • These genes are clustered on chromosome 3 in the homeotic complex (also called Hom-C) in two regions- • The Antennapedia complex- • The bithorax complex- Two amazing features 1. The order of these genes on the chromosome matches order of segmental expression Chromosome 3 2. Homeotic genes are regulated by all gene products expressed posterior to it
What about dorsal ventral polarity?? • This occurs after cells are created (post syncytial blastoderm) • Dorsal (a transcription factor) gradient is established • Dorsal is found throughout syncytial blastoderm, but only in nuclei of ventral cells How does this occur? By a very complex pathway involving gurkin and torpedo proteins ( and a host of other proteins) Organs form at the intersection of dorsal-ventral and anterior-posterior regions of gene expression