Download

1 / 68
680 likes | 685 Views
Learn about protein structure, types of prosthetic groups, protein purification methods, and enzyme kinetics with examples like hemoglobin and chemical reactions.

E N D
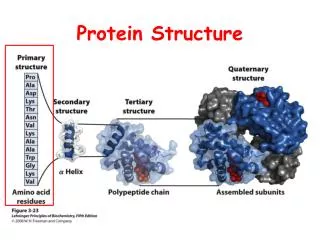
Last updated: 9/22/11 12:44 PM Lec. 6 Intro Bio 2011 Protein structure cont. Prosthetics groups Membrane proteins Protein purification methodsUltracentrifugation Native gel electrophoresis SDS gel electrophoresis Gel filtration Chemical kinetics Enzyme kinetics Michaelis-Menton equation turnover number, Km
Hemoglobin α2β2 tetramer Riboflavin~ vitamin B2 Heme Most prosthetic groups are bound tightly via ONLY weak bonds. Tetrahydrofolic acid ~ vitamin B9 heme
Metal ions can be prosthetic groups A “zinc finger” protein domain with two histidines and two cysteines shown Zinc (Zn++) (Voet and Voet, Biochemistry)
Membrane proteins Hydrophobic side chains on the protein exterior for the portion in contact with the interior of the phospholipid bilayer. Anions are negatively charged. Cations are positively charged
Small molecules bind with great specificity to pockets on protein surfaces Too far
Estrogen receptor binding estrogen, a steroid hormone detail estrogen estrogen Estrogen receptor is specific, does not bind testosterone
Protein binding can be very specific Testosterone Estrogen Estrogen (green) binding to amino acid side chains in the estrogen receptor
Protein separation methods Ultracentrifugation Mixture of proteins
Causing sedimentation: centrifugal force = mω2r m = mass ω = angular velocity r = distance from the center of rotation Opposing sedimentation = friction = foV. fo = frictional coefficient (shape) V = velocity Constant velocity is soon reached; then, no net force So now: centrifugal force = frictional force (balanced each other out) V = mω2r/fo, And so: mω2r = foV Or: V = [ω2r] x [m / fo]
12 Sample loaded here Glass plates Winner: Small, +++ High positive charge + + + Loser: Large, + low positive charge +++ +++ +++ poly- acrylamide fibers + Intermediate: Large, +++ high positive charge + + Intermediate: Small, + Low positive charge +++ +++ +++ Molecules shown after several hours of electrophoresis
Upper resevoir Upper resevoir Cut out for contactof buffer with gel Cut out for contactof buffer with gel
Cut out of glass platefor contact of buffer with gel Clamped glass sandwich Electrodeconnection(~ 150 V) Reservoir for buffer Platinum wire electrode
Power supply Happy post-doc Tracking dyes
SDS PAGE = SDS polyacrylamide gel electrophoresis sodium dodecyl sulfate, SDS (or SLS): CH3-(CH2)11- SO4-- CH3-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-SO4-- SDS Polypeptides denatured, act as random coils All have same charge per unit length All subject to the same electromotive force in the electric field Separation based on the sieving effect of the polyacrylamide gel Separation is by molecular weight only SDS does not break covalent bonds (i.e., disulfides) (but treat with mercaptoethanol for that, boil for a bit for good measure)
Reduction of protein disulfide bonds : Protein-CH2-S-S-CH2-Protein + 2 HO-CH2CH2-SH) (mercaptoethanol) [ a reducing agent ] Protein-CH2-SH+ HO-CH2CH2-S-S-CH2CH2-OH +HS-CH2-Protein Protein gets reduced; mercaptoethanol gets oxidized
e.g., “p53” Molecular weight markers (proteins of known molecular weight) P.A.G.E. 12 18 48 80 110 130 160 140 Kd
Molecular sieve chromatography (= gel filtration, Sephadex chromatography) Sephadex bead Mild conditions (pH, temperature), proteins in their native state
Molecular sieve chromatography Sephadex bead
Molecular sieve chromatography Sephadex bead
Molecular sieve chromatography Sephadex bead
Molecular sieve chromatography Sephadex bead
Plain Fancy 4oC (cold room)
Larger molecules get to the bottom faster, and …. Non-spherical molecules get to the bottom faster ~infrequent orientation Non-spherical molecules get to the bottom faster
Largest and most spherical Lowest MW Winners: Largest and least spherical Similar to handout 4-3, but Winners &native PAGE added Most chargedand smallest Winners:
Enzymes = protein catalysts
Each arrow = an ENZYME Each arrow = an ENZYME
H2 + I2 H2 + I2 2 HI 2 HI + energy Chemical reaction between 2 reactants “Spontaneous” reaction: Energy released Goes to the right H-I is more stable than H-H or I-I here i.e., the H-I bond is stronger, takes more energy to break it That’s why it “goes” to the right, i.e., it will end up with more products than reactants i.e., less tendency to go to the left, since the products are more stable
Atom pulled completely apart (a “thought” experiment) 2H + 2I say, 100 kcal/mole say, 103 kcal/mole Change in Energy (Free Energy) H2 + I2 { -3 kcal/mole 2 HI Reaction goes spontaneously to the right If energy change is negative: spontaneously to the right = exergonic: energy-releasing If energy change is positive: spontaneously to the left = endergonic: energy-requiring
H2 + I2 2 HI H2 + I2 2 HI H2 + I2 2 HI Different ways of writing chemical reactions H2 + I2 2 HI H2 + I2 2 HI
But: it is not necessary to break molecule down to its atoms in order to rearrange them 2H + 2I say, 100 kcal/mole say, 103 kcal/mole Change in Energy (Free Energy) H2 + I2 { -3 kcal/mole 2 HI
+ I I I I I I I I I I (H2 + I2) H H H H H H H H H H Transition state (TS) + (2 HI) Reactions proceed through a transition state + Products
2H + 2I ~100 kcal/mole Change in Energy H-H | | I-I (TS) Say, ~20 kcal/mole H2 + I2 Activation energy { -3 kcal/mole 2 HI
HHII (TS) Allows it to happen Energy needed to bring molecules together to form a TS complex Change in Energy (new scale) Activation energy determines speed = VELOCITY = rate of a reaction H2 + I2 { 3 kcal/mole 2 HI Net energy change: Which way it will end up. the DIRECTION of the reaction, independent of the rate 2 separate concepts
Direction We need it to go in the direction we want Speed We need it to go fast enough to have the cell double in one generation time Concerns about the cell’s chemical reactions
Example Biosynthesis of a fatty acid 3 glucose’s 18-carbon fatty acid Free energy change: ~ 300 kcal per mole of glucose used is REQUIRED So: 3 glucose 18-carbon fatty acid So getting a reaction to go in the direction you want is a major problem (to be discussed next time)
Direction We need it to go in the direction we want Speed We need it to go fast enough to have the cell double in one generation time Catalysts deal with this second problem, which we will now consider Concerns about the cell’s chemical reactions
The velocity problem is solved by catalysts The catalyzed reaction The catalyst takes part in the reaction, but it itself emerges unchanged
HHII (TS) Activation energy without catalyst TS complex with catalyst Change in Energy Activation energy WITH the catalyst H2 + I2 2 HI
Reactants in an enzyme-catalyzed reaction = “substrates”
Reactants (substrates) Active site or substrate binding site (not exactly synonymous, could be just part of the active site) Not a substrate
Substrate Binding Unlike inorganic catalysts, enzymes are specific
Small molecules bind with great specificity to pockets on ENZYME surfaces Too far
Unlike inorganic catalysts, enzymes are specific succinic dehydrogenase HOOC-HC=CH-COOH <-------------------------------> HOOC-CH2-CH2-COOH +2H fumaric acid succinic acid NOT a substrate for the enzyme: 1-hydroxy-butenoate: HO-CH=CH-COOH (simple OH instead of one of the carboxyls) Maleic acid Platinum will work with all of these, indiscriminantly
+ • Enzymes work as catalysts for two reasons: • They bind the substrates putting them in close proximity. • They participate in the reaction, weakening the covalent bonds • of a substrate by its interaction with their amino acid residue side groups (e.g., by stretching bonds).
Dihydrofolate reductase, the movie: FH2 + NADPH2 FH4 + NADP or: DHF + NADPH + H+ THF + NADP+ Enzyme-substrate interaction is often dynamic. The enzyme protein changes its 3-D structure upon binding the substrate. http://chem-faculty.ucsd.edu/kraut/dhfr.mpg
Substrate Product (reactants in enzyme catalyzed reactions are called substrates) S P Velocity = V = ΔP/ Δ t So V also = -ΔS/ Δt (disappearance) From the laws of mass action: ΔP/ Δt = - ΔS/ Δt = k1[S] – k2[P] For the INITIAL reaction, [P] is small and can be neglected: Initial ΔP/ Δt = - ΔS/ Δt = k1[S] So the INITIAL velocity Vo = k1[S] back reaction Chemical kinetics O signifies INITIAL velocity