Download

1 / 25
250 likes | 418 Views
Mutualism, Symbiosis, and the Serial Endosymbiosis Theory. Reading: Freeman, pps 615-617. Endosymbiosis.

E N D
Mutualism, Symbiosis, and the Serial Endosymbiosis Theory Reading: Freeman, pps 615-617
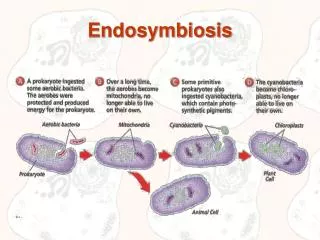
Endosymbiosis • Symbiosis, which means "living together" is an ecological interaction in which two or more species live in or on each other in close contact. Thus, endosymbiosis is when an organism actually lives within another organism.
Mutualistic symbioses are such interactions where the two parties benefit each other in terms of the ability of each partner to survive and reproduce. • Certain such mutualistic symbioses are some of the most important ecological interactions on the planet today. • For instance, the interaction between coral polyps (animal phylum cnidaria, class anthozoa) and their endosymbiotic zooxanthellae (mostly dinoflagellates, but also other photosynthetic partners) are essential to the productivity and long-term survival of coral reef ecosystems.
Likewise, the well-know partnership between legumes (plant family leguminaceae) and nitrogen-fixing bacteria (genera rhyzobium and bradyrhyzobium) is pivotal to the survival of terrestrial plant communities.
Termites (insect order isoptera) harbor endosymbiotic protists, without which they would not be able to digest wood, and in many tropical communities, termites are essential for the process of returning the nutrients locked in wood to the soil.
In hydrothermal vent systems, productivity is driven partially by chemoautotrophic archaeans living in an endosymbiotic relationship with tube worms (actually their own phylum, the vestimentaria, genus riftia).
Mutualistic endosymbioses between two partners are often complex associations that are regulated by the genetic interactions of the partners, and many are very ancient. • For instance, it is likely that the ecological interactions between corals and zooxanthellae go back to the Triassic period, at the beginning of the age of dinosaurs. Some endosymbiotic mutualisms are much older than that.
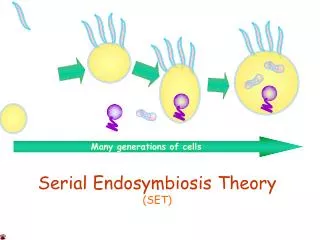
Endosymbioses are also thought to be important in an evolutionary context as well. • A theory for the evolution of subcellular organelles was postulated by Lynn Margulis in 1981. • A radical notion at the time, she postulated that mitochondria, chloroplasts, and possibly some other subcellular organelles, originated by the incorporation of a symbiotic organism into the eukaryotic genome. This is the Serial Endosymbiosis theory, SET. • SET, postulates that a series of endosymbiotic events enabled the evolution of eukaryotes, as evidenced by the DNA still present in mitochondria and chloroplasts. • Aspects of this theory are becoming increasingly well supported (by phylogenetic studies, and other evidence from subcellular ultrastructure), and it is possible that endosymbiosis is a major mechanism of macroevolution, allowing an organism to acquire major new functions by incorporating the genome of another lineage into itself.
Original Evidence in Favor of SET • The following observations let Lynn Margulis to propose SET. • Note that, since they were used to formulate the theory, they cannot be used to test it, and they do not exclude other explanations. • Mitochondria and chloroplasts are about the size of an average bacterium • Both organelles replicate by fission, like bacteria. Aditionally, the duplication of both organelles takes place independently of nuclear division. • Both organelles have their own ribosomes and manufacture their own proteins. These ribosomes resemble bacterial ribosomes, and are poisoned by streptomyacin, which does not affect eukaryote ribosomes. • The photosynthetic organelle of glaucophyte algae has an outer layer containing peptidoglycan-which is found in cyanobacteria but not normally found in eukaryotes. • Cyanobacteria have a similar structure to chloroplasts, and contain the same chlorophyls. • Mitochondria and chloroplasts have their own genomes-these are circular molecules resembling bacterial chromosomes.
Support for SET • Strong support for SET came from the field of evolutionary systematics. • Years after the theory was proposed, sequencing of the genomes of mitochondria demonstrated that, within the tree of life, they cluster where we would expect them to be if they evolved from proteobacteria. • Likewise, chloroplast genomes cluster near the cyanobacteria on the tree of life. • Neither of these observations would be expected if SET were incorrect.
The origin of chloroplasts or mitochondria is referred to as a primary endosymbiotic event. • Secondary endosymbiotic events occur when an organism that has already underwent endosymbiosis is incorporated into still another organism via endosymbiosis. • For instance, it is probable that green and red algae acquired photosynthesis via a primary endosymbiosis with cyanobacteria. • Very likely, other groups of protozoa have acquired photosynthesis via secondary endosymbisis with either • green algae (euglenoids, such as Euglena arcus) • red algae (diatoms, certain dinoflagellates, and others) • or both. Some groups are even tertiary endosymbiotes.
Compare the generalized prokaryotic (top) and eukaryotic (bottom) cells. The eukaryote is defined as having a membrane bound nucleus, whereas the prokaryote does not. • When you compare this eukaryotic cell to the animal cell, note that the typical animal cell contains fewer components than this eukaryotic cell; that is because this cell is an amalgam of four diverse kingdoms of organisms, whereas the animal cell demonstration represents only one.
Evolutionary Time • This diagram shows three major endosymbiotic events in eukaryotic evolution. • The grey branch represents the Prokaryotic genome (Kingdom Monera). • The white branch represents the Eukaryotic genome; it is not rooted because originated from the prokaryotic genome by endosymbiosis.
Prokaryotes,Kingdom Monera, populated the planet for years long before any eukaryotic life existed. Combinations of different prokaryotic genomes by endosymbiotic events created the diversity of more complex life that exists today. • The Animals feature a eukaryotic cell with mitochondria but no plastids. Animals must consume sugars produced by photosynthetic organisms in order to survive. they have fluidic cell walls and produce haploid gametes. • The Fungi are composed of eukaryotic cells containing mitochondria but no plastids. Their cell walls, reproductive methods, and nutrient requirements differ significantly from animal cells, but moreso from plant cells. • The Protists, more correctly dubbed protoctists, are unicellular organisms; they are eukaryotic cells that possess either both mitochondria and chloroplasts (upper lineage) or mitochondria only ( lower lineage). • The Plants have all three endosymbiotic events: a eukaryotic cell with mitochondria and chloroplasts. They have rigid cell walls and vacuoles, create polyploidy gametes, and produce sugars. • Archaeans, or archaeabacteria, inhabit some of the most harsh environments on Earth. Their cytoplasm and genetic arrangements make certain archaeans good candidates for the nucleocytoplasm of the eukaryotic cell.
A chloroplast is a type of plastid; possibly, there are many plastids that have become endosymbionts, leading to a vast array of photosynthetic organisms. • The chloroplast, ubiquitous to the green land plants, was derived of a cyanobacterium
The chloroplast is evolutionarily derived from a cyanobacterium that was phagocytized by a hungry eukaryotic ancestor. • Photosynthetic reactions in the chloroplast occur in and between the thylakoid membranes, which are stacked into grana. • Compared to free-living blue-green algae, the chloroplast has far more thylakoids, indicating the increased amount of work that the chloroplast must do to support an entire cell rather than just itself.
Spirochaetes, a group of highly motile cork-screw-like bacteria, are thought to have become undulipodia, kinetosomes, and basal bodies-- the constituents of a eukaryotic cell that confer movement and aid in mitotic division. • The arrangement of microtubules in these organelles looks very much like the structure of a spirochaete. • Spirochaetes feed by attaching to a host, much like a leech; in this way one can visualize attachment leading to a symbiotic union. • The lack of genetic evidence (thusfar) linking spirochaetes and organelles, however, spirochaetes remain an unaccepted and controversial part of SET.
Mitochondria • Although not all eukaryotes contain mitochondria, the majority of them do. • According to SET, mitochondria were acquired by a symbiotic event. • It is still uncertain what prokaryote the mitochondrial ancestor was, but their genomes resemble those of proteobacteria. • We can speculate about the details of its structure.
The Mitochondrion: A Metabolic Acquisition • In the changing atmosphere of the early earth, oxygen was toxic, a metabolic waste product accumulating in the atmosphere. Life on the surface of the planet was a battle of novel metabolic strategies: organisms that became able to tolerate and eventually metabolize oxygen had a selective advantage. • When the ancestor of modern-day mitochondria-containing eukaryotes acquired an endosymbiont that used oxygen to metabolize sugars with a high energy yield, it gained this evolutionary “edge.” The abundance of eukaryotic life exploiting this metabolic advantage today is a tribute to the potential impact of endosymbiotic events.
Inside a Mitochondrion • The mitochondrion is organized into two compartments by a double membrane system. • The outer membrane is highly porous, with open channels for any molecule less than 10 kiloDaltons (the size of the smallest proteins). As a result, ADP, ATP, sugar molecules, and many other molecules integral to mitochondrial functions can pass into the inter-membrane space. • The inter-membrane space is a crucial part of oxidative metabolism. In conjunction with the mitochondrial matrix, a proton gradient can be established as protons are pumped into the inter-membrane space during the reactions of oxidative metabolism. • The inner membrane contains the host of the proteins that are the reaction centers for oxidative metabolism. This membrane is MUCH harder to get through than the outer membrane, being only freely permeable to carbon dioxide, oxygen, and water, and otherwise regulated by protein channels. The folds of the membrane, or cristae, dramatically increase the surface area of the inner membrane and therefore increase the amount of energy that can be produced. • The mitochondrial matrix has the components of its free-living ancestry that the mitochondrion still uses, such as its genetic material and ribosomes. The matrix also contains the components necessary to perform the reactions of the Citric Acid Cycle, the portion of respiration whose products are the fuel for oxidative metabolism.
Oxidative metabolism is an electron-transporting set of reactions that occur on between the mitochondrial membranes. • The Kreb's cycle, where small energy carrying molecules are produced from processing sugar, occurs in the matrix provide the energy required to drive the reactions on the inner membrane (IM). • The complexes in the inner membrane transport electrons, but not protons, so protons liberated from the energy-transporting molecules are are actively pumped into the inter-membrane-space (IMS) while the electrons are pushed along in a transport chain. • Oxygen is the final electron acceptor, yielding water and terminating the chain. • The last complex in the process, ATP synthase, uses the emergent proton gradient to produces ATP, a very high-energy molecule usable by the rest of the body.
The Phagocytotic Theory of Mitochondrial Acquisition • Tare two theories of mitochondrial acquisition: phagocytosis and predation. • In the phagocytotic model, an Archaean envelops a purple sulfur-loving eubacterium. The Eubacterium, instead of being digested, persists inside the cell. • Host • The host that acquired the first mitochondrion-bound endosymbiont donated the nucleocytoplasm to the eukaryotic cell. • This host would have been • microaerophilic (tolerating a very small quantity of air) • a sulfur-reducing metabolism, • lacking a cell wall (being therefore capable of phagocytosis) • containing genetic material organized by histone-like proteins. • An evolutionary event in which the archaean acquired or developed the cytoskeletal structure required for phagocytotic activity would have had to have occurred previously. • A combination of the archaeans (or archaeabacteria) Thermoplasma (for its lack of a cell wall and histone-like proteins) and Sulfolobus (for its metabolic pathways) is shown here to reconstruct what the common ancestral host may have looked like: it is microaerophilic, wall-less, and has arginine and lysine-rich histonelike proteins that coil and protect its genetic material from acid and heat shock in the extreme environments that it inhabits. • Neither Thermoplasma nor Sulfolobus, however, is genetically similar enough to a modern-day eukaryote to actually be the common ancestor.
12 Phagocytotic Theory: Endosymbiont • The prokaryotic endosymbiont in the phagocytotic theory would have been capable of a very similar oxygen-based metabolism to modern day mitochondria, and is represented here based on the structure of the micrococcus Paracoccus. • Paracoccus is a good model because it has cytochromes and ubiquinones in its electron transport chain that look like those in the oxidative metabolism of mitochondria. • Although Paracoccus does not genetically resemble a mitochondrion outside of its respiratory metabolism, other alpha-proteobacteria have genetic sequences that more closely resemble present-day mitochondria."
The Predatory Theory • In contrast to the phagocytotic theory, in the predatory theory, the endosymbiont is the aggressor. • The predatory prokaryote attaches to and subsequently burrows through the outer layer of the host's plasma membrane, eventually inhabiting the periplasmic space. • Once inside the periplasmic space, the predator's flagellum disappears, and, under normal conditions, it begins to digest the host. • In this case, however, the aggressor fails to destroy its prey and persists inside the host as an endosymbiont. Bdellovibrio life cycle
According to this theory, the host would have been pretty much the same as in the phagocytotic theory • The prokaryotic endosymbiont would have been • capable of a very similar oxygen-based metabolism to modern day mitochondria • capable of this invasive action, and is represented here based on the structure of the pseudomonad Bdellovibrio. • Bdellovibrio is a good structural candidate for the common ancestor of the endosymbiont because it is aerobic and invades the periplasmic space, feeding on cellular material, and stays there to reproduce. • A failed attempt at this invasive strategy would leave the endosymbiont surrounded by a host-derived membrane: the same result as in the phagocytotic model. • Although Bdellovibrio does not genetically resemble a mitochondrion and, although aerobic, does not have a metabolism that looks as much like mitochondrion metabolism as Paracoccus does, other alpha-proteobacteria have genetic sequences that more closely resemble present-day mitochondria.