Download

1 / 23
240 likes | 523 Views
Section 7. Lipid Metabolism. Fats: fatty acid biosynthesis. 11/04/05. Oxidation of Fatty Acids Other Than Palmitate. Any even number of saturated carbons does not require any additional enzymes. Products are as for palmitate.

E N D
Section 7. Lipid Metabolism Fats: fatty acid biosynthesis 11/04/05
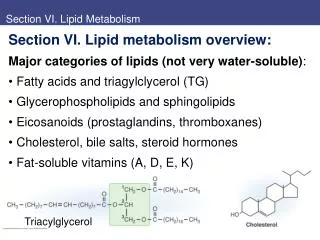
Oxidation of Fatty Acids Other Than Palmitate • Any even number of saturated carbons does not require any additional enzymes. Products are as for palmitate. • An odd number of saturated carbons does not require any additional enzymes. Same products plus one propionyl CoA. • Unsaturated fatty acids require additional enzymes. Same products, but less energy, compared to saturated fatty acid the same length. • See table 12.1 for a list of common fatty acids. 1
An odd-numbered double bond moves into the 3-position, which is not a substrate for the principal enzymes of the cycle. An isomerase moves it to the 2-position. Moving the double bond is energy neutral, but one less FADH2 is made because one acyl CoA dehydrogenase step is “skipped.” As acetyl CoA’s are removed from an unsaturated fatty acid, double bonds move into or near the active sites of the -oxidation cycle enzymes. Double Bonds in Odd-Numbered Positions 16:1 cis-D9 (p 610) 2
Double Bonds in Even- Numbered Positions. • An even-numbered double bond moves to the 4-position. • Acyl CoA dehydrogenase oxidizes it producing FADH2 and the non-substrate dienoyl CoA. • The additional enzyme 2,4-dienoyl CoA reductase uses NADPH to produce a position double bond. • The additional enzyme cis D3-enoyl CoA isomerase (see above) moves the double bond from the 3- to the 2-position. • NADH is produced after hydration, and then acetyl CoA, as usual (not shown). • The net effect for the cycle is the equivalent of one less NADH. 3
Fig. 22.10 modified Example: Linoleoyl CoA C18:2 cis-D9,cis-D12 + 3 acetyl CoA + 3 FADH2 + 3 NADH + acetyl CoA + NADH (no FADH2) 4
Production is enhanced by low carbohydrate (diabetes, starvation) and/or low O2 (hypoventilation) General anesthesia: CO2 up, pH down, ketone bodies up. Volatile acetone formation is non-enzymatic. The liver normally converts acetyl CoA to ketone bodies that are used by peripheral tissues. See fig. 22.19 Ketone Bodies 5
Acetoacetate Utilization • In peripheral tissues, the ketone body acetoacetate is activated, and converted back to acetyl CoA. • 3-hydroxybutyrate and acetoacetate are favored over glucose by the renal cortex and cardiac muscle. Fig. 22.20 6
Summary of Fatty Acid Biosynthesis • When the cell energy level is high, rather than being used by the Krebs cycle, acetyl CoA is transferred from the mitochondrial matrix to the cytosol. • In the cytosol, acetyl CoA is converted to malonyl CoA, which is used by fatty acyl synthase (FAS) for the synthesis of palmitate. • Palmitate is transported to adipose tissue and used to synthesize triacylglycerol. • The palmitate synthetic reactions are reversals of the degradative reactions, but the enzymes, cofactors and locations are different. 7
Reactions on the right are catalyzed by FAS in the cytosol. Compare degradation and synthesis structures. Fig. 22.2 8
Citrate Shuttle Transfers Acetyls to Cytosol • High [ATP] inhibits the Krebs cycle; [citrate] increases. • Citrate translocase enables citrate and pyruvate to cross the mitochondrial inner membrane. CoA does not cross (remember acyl CoA / acyl carnitine). • NADPH is made at the expense of NADH in the cytosol. ATP + CO2 Fig. 22.25 +CO2 9
In the cytosol, acetyl CoA is carboxylated to make the activated precursor, malonyl CoA. This is the committed step in fatty acid biosynthesis. ATP provides energy. Biotin is a cofactor. Two sequential reactions occur in the active site. Activation by Acetyl CoA Carboxylase (p 617) (p 618) biotin-Enz + ATP + HCO3- CO2~biotin-Enz + ADP + Pi CO2~biotin-Enz + acetyl CoA malonyl CoA + biotin-Enz 10
Biotin: a CO2 Carrier • ATP reacts first providing energy to bind and activate HCO3-. • Next acetyl CoA binds and the activated CO2- is transferred to the acetyl group. Figs. 24-10 and 24-11 (Stryer 4th) 11
Fatty Acid Synthase Reactions CE Condensation forms 4 carbon unit on acyl carrier protein (ACP). Reduction of ketone to hydroxyl by NADPH. (KR) CE + Fig. 22.22 12
Fatty Acid Synthase Reactions, con’t • Dehydration produces a double bond. • Reduction to a saturated 4 carbon fatty acid chain. (DH) (ER) Fig. 22.22 13
FAS is a Dimer • Malonyl transfer (MT), acetyl transfer (ATP and condensation (CE) on one subunit. • Reduction (KR), dehydration (DH), reduction (ER) and thiolysis (TE) on the other subunit. • The growing FA chain is passed between subunits by ACP. Fig. 22.23 14
Acyl carrier protein (ACP) • ACP has a long flexible chain, derived from pantothenic acid, to which the growing fatty acid is attached. Fig. 22.21 15
Condensation • Both subunits of FAS are involved. • Condensation is catalyzed by CE on upper subunit. Fig. 22.24 16
Reduction • 2 NADPH are used. • Reduction (reduction, dehydration, reduction see slides 12 &13) occur on lower subunit of the dimer. Fig. 22.24 17
Translocation and binding of a new malonyl CoA • The 4 carbon chain is transferred to CE. • A new malonyl CoA binds ACP on other subunit. • The cycle (condendation, reduction, dehydration, reduction) repeats until 16 carbon palmitate is formed (not shown). • Palmitate is released by TE (see slide 14). Fig. 22.24 18
Net Reaction for Palmitate Synthesis 8 acetyl CoA + 7 ATP + 14 NADPH + 6 H+ palmitate + 14 NADP+ + 8 CoA + 6 H2O + 7 ADP + 7Pi For this reaction, there are 7 (for 7 ATP) + 35 (for 14 NADPH) = 42 ~P equivalents used. Compare to 26 obtained from the palmitate conversion to acetyl CoA by fatty acyl CoA synthetase and the b-oxidation cycle. (p 622) 19
Control of Fatty Acid Synthesis is at the Committed Step • Fig. 24-18 Styer 4th 20
Control of Acetyl CoA Carboxylase Activity • Phosphorylation by kinase inhibits carboxylase (+AMP, -ATP). • Phosphatase activates carboxylase (+insulin, -glucagon, & epinephrine). • Citrate partially activates the inactive phosphorylated acetyl Co A carboxylase allosterically. • Palmitoyl CoA inhibits carboxylase and citrate translocase. Fig. 22.26 21
Web links Odd Chain Fatty Acids. The fate of propionyl CoA. Unsaturated Fatty Acid Oxidation. The role of isomerase. Next Topic: Membrane lipids.