Download

1 / 36
361 likes | 601 Views
Sensory Encoding of Smell in the Olfactory System of MAMMALS (reviewing “Olfactory Perception: Receptors, Cells, and Circuits” by Su et al, 2009) Ben Cipollini COGS 160 May 13, 2010. TODAY. Compare / contrast! Gross Pathways Receptor Neurons Glomeruli Output Neurons Higher centers.

E N D
Sensory Encoding of Smell in the Olfactory System of MAMMALS (reviewing “Olfactory Perception: Receptors, Cells, and Circuits” by Su et al, 2009) Ben Cipollini COGS 160 May 13, 2010
TODAY • Compare / contrast! • Gross Pathways • Receptor Neurons • Glomeruli • Output Neurons • Higher centers
Gross Pathways • ORNs in antennae • Projection neurons from antenna lobe to lateral horn and mushroom body • Glomeruli in antenna lobe mediate most receptor-specific processing • Kenyon cellsin mushroom body have sparse representation of odors for associative learning • Lateral horn has place-specific processing of sensory-motor associations Keene & Waddel (2007)
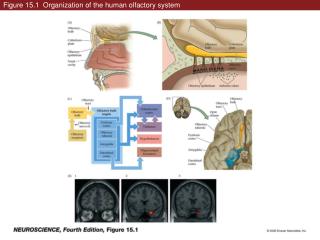
Insects: 2 organs Antennae Maxillary Palps Olfactory Organs • Mammals: 4 organs • Main Olfactory Epithelium • Vomeronasal Organ • Grueneberg ganglion • Septal organ of Masera
Main Olfactory Epithelium • MOE • ORs for odor ID; 250-1200 functional genes • Trace amine-associated receptors (TAARs) can detect volatile urine-based amines; 15 in mouse (social cues) • Output to main olfactory bulb X X X
Vomeronasal Organ • V1Rs (urine) for conspecific recognition, male sexual behavior, maternal aggression, regulation of female estrous cycles, stress level indicator • V2Rs (sweat and urine) for pregnancy blocking, individual / gender identity, aggression (from males), stress (from females) • Formyl Peptide Receptors (immune system) for health status • Outputs to accessory olfactory bulb
Gruenberg Ganglion • Trace amine-associated receptors (TAARs) • ONE V2R receptor • Responsive to mechanical stimulation (sniffing / air puffs) • Outputs to main olfactory bulb X
Septal Organ of Masera • ORs for general alerting • Responsive to mechanical stimulation (sniffing / air puffs) • Outputs to main olfactory bulb X
Antennae • Basiconic for odor recognition, repulsion behavior • 60-340 Ors • A few Grs (CO2) • Coleoconic (function unknown) • Ionotropic receptors → derived from glutamate receptors! • Trichoid for pheromones Keene & Waddel (2007)
Maxillary Palps • Basiconic sensillia for taste enhancement Keene & Waddel (2007)
The Evolutionary Story • Insects • Finding homologies in species of the same order can be challenging • Probably fast evolution • Mechanism (duplication & variation vs. modification) unknown • NOTE: loss of a single OR doesn't necessarily eliminate associated behavior • Ensemble encoding • Different ORs coding for an odor at different concentrations (helps with variable gain)
Review: Tuning Curves of ORNs • Odorants are identified by the pattern of receptors activated • Including inhibition of tonic firing • Individual receptors are activated by subsets of odorants • Receptors lie along a smooth continuum of tuning breadths • Broadly tuned receptors are most sensitive to structurally similar odorants • Higher concentrations of odorants elicit activity from greater numbers of receptors • Odor intensity as well as odor identity is represented by the number of activated receptors Hallem et al (2006)
ORN Activity vs Concentration Kreher et al (2008)
ORN Activity vs Concentration Kreher et al (2008)
Aquatic vertebrates Only have OR class I Terrestrial vertebrates Have OR class I & II Dolphins Have class I 100% of OR class II pseudogenized Evolution II: Pseudogenization • Humans • 20-30% of ORs • 90% of VRN1s • 100% (so far) of VRN2s (only 20 genes exist)
For no particular reason... • 3 cool properties of ORNs that were discussed in this paper: • Temporal tuning curves • Antagonistic ORNs! • Insect ORNs are actually really weird!
Tuning Curves of ORNs (New):Temporal Dynamics • Different ORNs can have different temporal dynamics (even for the same odor) • A single ORN can have different temporal dynamics to different odors Odorant tuning curves Bruyne et al (2001)
Combinatorics: Antagonistic Inhibition • The perceived magnitude of an odorant mixture was neither additive nor a simple average of its components • Fell between these limits, due to: • Masking (i.e. modification of perceived odor) or counteraction (i.e. reduction of odor intensity). • Mixing some odorants led to the emergence of novel perceptual qualities that were not present in each individual odorant • Suggests that odorant mixture interactions occurred at some levels in the olfactory system • Observed at presynaptic ORN axons in olfactory bulb Oka et al (2004)
Insects ORNs are CRAAAZY! • Insect odor receptors have 7 transmembrane domains and have long been assumed to be GPCRs. • BUT we see major major differences! • No G protein mutant has been found to suffer a severe loss of olfactory function. • The topology of the insect Ors is inverted relative to GPCRs. • Each OR also appears to form a heteromultimer with Or83b • A canonical OR (with Or83b), can form a “ligand-gated cation channel” • Due to an odorant-induced, rapidly developing, transient inward current, independent of G protein signaling • A second, slower and larger component to the odorant-induced inward current • Slower both in onset and decay kinetics • Is sensitive to inhibition by a GDP analog Siegel et al (1999)
ORN Transduction: “canonical” • Odorant binds to the odor receptor • Odor receptor changes shape and binds/activates an “olfactory-type” G protein • G protein activates the lyase - adenylate cyclase (LAC) • LAC converts ATP into cAMP • cAMP opens cyclic nucleotide-gated ion channels • Calcium and sodium ions to enter into the cell, depolarizing the ORN • Calcium-dependent chlorine channels contribute to depolarization as well • G protein turned off by GDP Firestein & Menini (1999)
Review: Projection Neurons • Live in antenna lobe (~200 per) • Receive input from ALL ORNs of a single class(~50; ~25 from each side) • Despite convergent input, show broader odorant tuning than ORNs • Project out to “higher centers”: mushroom body & lateral horn
Tufted & Mitral Cells • Live in olfactory bulb • Receive input from ALL ORNs of a single class from a single side • Like insect projection neurons, show broader odorant tuning than ORNs • Like insect projection neurons, project out to “higher centers” • NOTE: only mitral cells project to posterior piriform cortex
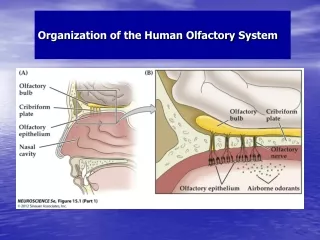
Review: Glomeruli • In Antenna Lobe, one per odorant “class” (50) • Consist of: • Axons of ORNs • Dendrites of projection neurons • Neurites (axons and dendrites) of local neurons • ORN inputs all from same “class”, come bilaterally Mammalian Glomeruli Kandel, Jessel, Schwartz (2000)
5000:1 convergence M/T input from 1 Interglomerular inhibition (granule cells) Intraglomerular inhibition (juxtaglomerular cells) Glomeruli: MAMMALIAN INSECT • 50:1 convergence • Pns input from 1 • Interglomerular inhibition (local neurons) • Intraglomerular inhibition (local neurons)
Review: Transformations • Two glomerular transformations: • Increasing signal-to-noise • Producing variable gain • PN / Kenyon Cell Transformations: • Decorrelation of ORN signals
Variable Gain Revisited • Broader tuning widths and nonlinear amplification among projection neurons are mainly due to strong ORN-projection neuron synapses How? • Low Activity Amplification: Weak presynaptic ORN activity is sufficient to trigger robust neurotransmitter release and cause substantial PN responses. • High Activity Fall-off: Strong ORN activity leads to depletion of synaptic neurotransmitter. How about mammals? • The strong synapses : due to presence of numerous synaptic vesicle release sites and a high release probability • High probabilities of vesicle release have also been found in the mammalian olfactory bulb
Sparse Coding in Kenyon Cells • IN THE LOCUST • PNs (columns) respond to most odorants; KCs (columns) respond to very few • “Population sparseness” - % of cells that do NOT respond to an odor (rows) Perez-Orive et al (2002)
How Do Locust KCs Become Sparse? • High convergence (400:1, 50% of PNs!) • Weak unitary synaptic connections • Synaptic integration in (oscillatory) time windows • Voltage-gated channels amplify coincident spikes • High spiking threshold (50-100 coincident Pns) • Loss of oscillations in bees → no “fine” discriminations Fig. 7 from Masse et al (2009)
Higher-level pathways • Posterior Piriform cortex does classification (like Mushroom body!) • Cells within PPC project to MOST areas that are connected to • This includes feedback projections to olfactory bulb Johnson et al (2000)
Olfactory Learning Li et al (2008)