Download
1 / 17
170 likes | 246 Views
Perception Chapter 4 Visual Process beyond the Retina.

E N D
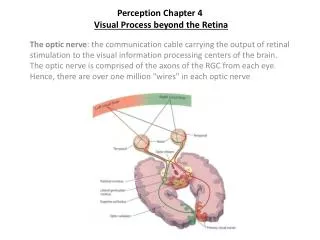
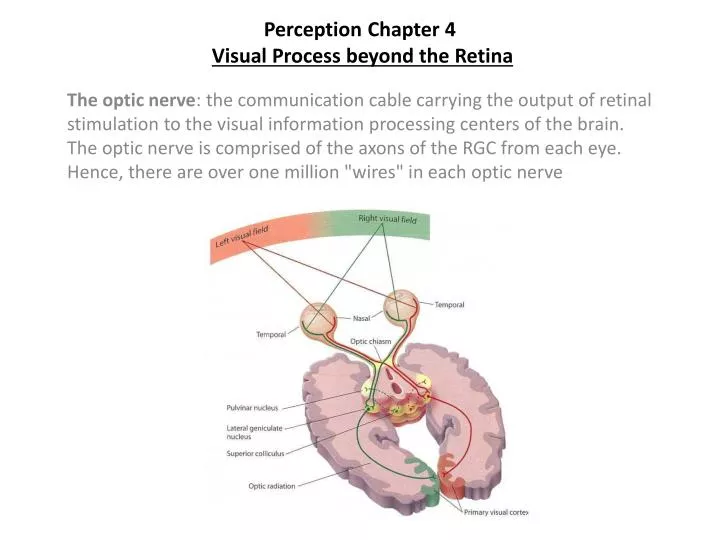
Perception Chapter 4Visual Process beyond the Retina The optic nerve: the communication cable carrying the output of retinal stimulation to the visual information processing centers of the brain. The optic nerve is comprised of the axons of the RGC from each eye. Hence, there are over one million "wires" in each optic nerve
Perception Outline #5Visual Process beyond the Retina a) optic chiasm: crossover point where fibers from two eyes meet b) ipsilateral fibers: the half of the optic nerve from each eye emenating from the temporal half of each retina, which do not cross at the optic chiasm c) contra-lateral fibers: the other half of the optic nerve, emanating from the nasal half of each retina, which do cross. d) optic tracts: term referring to the post-chiasm optic nerve, this represents combined fibers from both eyes. Why do half the fibers cross? Ans: Nasal half of one eye, and temporal half of other eye, monitor same visual field, therefore, fibers emanating from same areas monitoring same physical space are combined. About 80% of these fibers will project to the Lateral Geniculate Nucleus of the thalamus, the other 20% will project to a structure in the mid-brain called the Superior Colliculus.
Visual info processing in the Superior Colliculus: SC is the more phylogenticallyprimitive projection site, in some creatures such as fish and frogs, SC is major site of visual info processing. 1) Cells in SC have ill-defined "On" "Off" regions, therefore will respond to most any visual stimulus regardless of shape, color, or orientation. 2) SC cells seem most interested in location of visual stimulus, not identity of visual stimulus, therefore, SC has often been referred to as the "where" system, rather than the "what" system. 3) SC appears to have important role in directing ballistic eye movements toward a visual stimulus in the periphery 4) Most of the fibers projecting to SC are from the M type of RGC, therefore, messages often get to SC more quickly than to LGN. So what does all this mean? Ans: it appears that SC is very important for regulating visually guided reflexive behaviours, for example, the SC would quickly signal the presence of an object coming toward the head from the visual periphery. SC only cares that there is something there, LGN will figure later what it was!
Visual info processing in LGN: Structure: 1) There are 6 distinct layers in LGN, first two layers have large bodied cells, these are called the magnocellularlayers, the next four layers have smaller bodied cells and are called the parvocellular layers. 2) Contra-lateral fibers from the nasal retina of one eye contact with layers 1, 4, and 6, while the ipsilateralfibers from the temporal retina of the other eye make contact with layers 2, 3, and 5. The same arrangement is true for each LGN. 3) Input from both eyes is "mapped" onto LGN layers in such a way as to preserve the retinal topography. Retinotopic maps of LGN.
LGN: Retinotopic maps Spatial layout on retina is preserved when transmitted to layers of LGN
Visual info processing in LGN: Function: 1) LGN contains cells with similar receptive field characteristics as RGC, except that "off" portions of cells exert even stronger inhibitory affect. This serves to accentuate to an even greater extent the "edges" and "borders" already identified by RGCs. 2) Parvocellular layers contain cells with receptive fields which respond differentially to color. These are called color opponent cells, most are red/green or blue/yellow color opponent. These cells receive most of their input from the P type of RGC. 3) Magnocellular layers are color blind and receive input mostly from M type of RGCs. 4) Both Magnocellular cells and Parvocellular cells can signal movement, but Magno cells respond to fast movement, Parvo respond to slow movement. 5) LGN may also receive "top-down" visual processing biases due to its interconnections with visual cortex. 6) Interconnections with RAS may serve as “volume control” for visual inputs.
Visual info processing in the Primary Visual Cortex: • PVC: the area in the very back of the occipital lobe which first receives info from connections to LGN, also known as "striate cortex," "area 17,“ or V1
Visual info processing in the Primary Visual Cortex: Like RGCs and cells of LGN, cells of the PVC have receptive fields which monitor a restricted zone in the retina, however, the number of cells responding to input from the foveal area of retina far exceeds the number responding to input from more peripheral regions. This is known as cortical magnification. • Cortical magnification: the disproportionally large amount of cortical tissue dedicated to processing the foveal retina compared to the peripheral retina. • Cortical magnification stands to reason if you consider the number of RGCs connected to photoreceptors in fovea vs. RGCs connected to photoreceptors in periphery. Remember degree of convergence?
Functional properties of cortical cells: Orientation specificity: cells in PVC differentially respond to stimuli of varying orientations. This was not true of LGN cells or RGC cells, both of which contain circular receptive fields insensitive to orientation. Receptive fields in PVC cells tend to be more oblong shaped. Simple cells: Well defined excitatory region; location important. Complex cells: less well defined excitatory region; location less important. Hyper-complex: length of contour important oblique effect
Functional properties of cortical cells: • Direction specificity: cells in PVC also differentially respond to movement in different directions. One cell may respond vigorously if a visual stimulus moves from left to right across the receptive field, but not respond as much if movement is from right to left, other cells do just the opposite(also up/down). Simple cells: slower movement. Complex cells: faster movement
Functional properties of cortical cells: • Binocular cells: Two receptive fields, one for each eye. Stronger response to one eye (ocular dominance). Cells give a most vigorous response when a stimulus of the same size, shape, and orientation is located in a particular position in 3-d space across the two retinae. Cells are important for the the visual system's use of the depth cue called retinal disparity (more on that later).
Visual cortex organization: Columns and hypercolumns • Cell columns: in the PVC cells (s,c,hc) are arranged in layers by their preferred orientation. • Cell hypercolumns: the layers of orientation specific cells are arranged such that an orderly incrementing of orientation change exists across layers until a complete cycle has been achieve. Include both eyes. • Layer 4: input layer, monocular cells • Blobs: non-orientation specific cells for color processing
Cortical cells as feature detectors • "Families" of cortical cells as feature detectors: the responding of any single cell may be ambiguous as to the true nature of the visual stimulus. For example, a vertical preferring cell may start to increase its responding as a stimulus approaches a more vertical orientation, but does that mean the stimulus is approaching vertical from a clockwise or counterclockwise position? Don’t know just from the responding of that one cell (ambiguity problem), however, the responding of another cell who prefers certain types of oblique orientations may clear up the ambiguity. This points to the importance of cells working as aggraded sets in order to process visual info.
Two visual pathways beyond the PVCDorsal: “where” or “how” pathway Ventral: “what” pathway
M vs. P pathways and Dorsal/Ventral pathways Seeing vs. Acting distinction. Segregation but not complete separation.