Download

1 / 60
0 likes | 17 Views
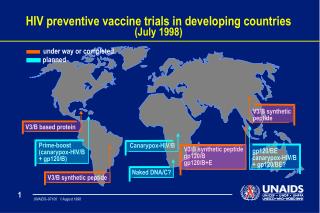
Developing a vaccine against HIV has proven to be extremely difficult due to the virus's ability to evade the immune system, rapid mutations, and diverse transmission mechanisms. Current vaccines struggle to stimulate broadly neutralizing antibodies, and the complexity of the virus itself poses a significant challenge. Various vaccine trials have shown minimal protection, highlighting the arduous and costly process of HIV vaccine development.

E N D