Download
1 / 1
10 likes | 234 Views
Characterization of exaggerated pain behavior and glial activation in a novel rat model of spinal cord injury A. L. Ellis 1 , J. Wieseler 1 , K. Brown 1 , C. Blackwood 1 , K. Ramos 1 , C. Starnes 2 , S. F. Maier 1 , L. R. Watkins 1 , & S. P. Falci 2

E N D
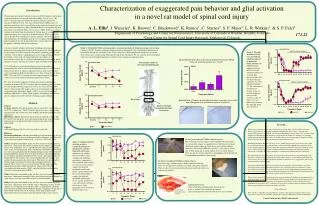
Characterization of exaggerated pain behavior and glial activation in a novel rat model of spinal cord injury A. L. Ellis1, J. Wieseler1, K. Brown1, C. Blackwood1, K. Ramos1, C. Starnes2, S. F. Maier1, L. R. Watkins1, & S. P. Falci2 1Department of Psychology and Center for Neuroscience, University of Colorado at Boulder, Boulder, Colorado 2Craig Center for Spinal Cord Injury Research, Englewood, Colorado Introduction Chronic pain after spinal cord injury occurs in 65% of patients and the pain is often described as severe and intolerable (Ridet, Trends Neuro., 20, 1997). Currently this pain reported by spinal cord injury patients is extremely difficult to treat. One successful method to reduce pain is to perform a dorsal root entry zone ablation, using neuroelectrical activity data along with transcutaneous c-fiber stimulation data to very specifically map the region to be lesioned (Falci, J Neurosurgey, 2002). In the past, studies looking at the underlying mechanisms of chronic pain associated with spinal cord injury have focused on neuronal changes. More recently the role of glia has been considered as having a primary role in the inception and maintenance of neuropathic pain in cases of spinal cord injury. Better understanding of the role of glia will lead us to more effective treatments for patients suffering with spinal cord injury. Current rat models of spinal cord injury including contusion and hemisection are problematic because of the negative side effects produced by the surgery itself. Rats often develop urinary retention, paralysis, and autotomy, which makes it challenging to assess exaggerated pain. We have developed a novel model of spinal cord injury that allows us to behaviorally evaluate the rats without compromising their health or motor function as well as allowing us to look at glial activation in the spinal cord. AV411 (Ibudilast) is a therapeutic drug used in varied models of neuropathic pain. AV411 crosses the blood-brain barrier, is well tolerated, and has been shown to suppress glial activation as well as prevent and reverse mechanical allodynia (Ledeboer et al., Expert Opin Investig Drugs, 2007). We used AV411 to explore whether suppressing glial activation could reverse the mechanical allodynia induced by avulsion. We show here the development of bilateral mechanical allodynia in response to unilateral T13/L1 avulsion, and corresponding glial activation over the month following surgery. In study 2, it is clear that our model shows a behavioral profile unique relative to the commonly used model of exaggerated pain, rhizotomy. Finally, AV411 reverses the mechanical allodynia induced by avulsion. Taken together, our findings indicate that glia are significantly involved in the resulting exaggerated pain. Methods Surgery Avulsion surgery- The dorsal spinal cord was exposed by laminectomy at the T13/L1 spinal level. The T13 and L1 dorsal roots were each isolated, clamped at the dorsal root entry zone (DREZ) and briskly pulled out of the spinal cord. Rhizotomy surgery- The dorsal spinal cord was exposed as above. The T13 and L1 dorsal rootlets were each isolated and gently transected with microscissors, leaving the proximal terminals in the spinal cord undisturbed. Subjects Adult male Sprague Dawley rats were used in each study. Behavior Mechanical allodynia- Absolute thresholds were determined using the up-down method with a series of calibrated von Frey filaments (von Frey test), as previously described (Chaplan et al., J. Neurosci. Methods, 1994). Study 1- Response thresholds on the von Frey test were recorded prior to and across a timecourse after either avulsion or sham surgery. Spinal cord tissues were collected 7, 14, 21, and 30 days post-surgery (n=5 avulsion and 1 sham at each timepoint). After overdosing with sodium pentobarbital, rats were transcardially perfused with physiological saline followed by 4% paraformaldehyde. Spinal cords were post-fixed in 4% paraformaldehyde for 15 minutes and then transferred to 30% sucrose. The spinal cords were blocked into 5-mm sections, one section including the site of injury and another 10-mm caudal to the site of injury. This latter site corresponds to L5/L6 which is the spinal site from which allodynia behaviors were elicited. These tissues were cryostat sectioned (20-um) and reacted by immunohistochemistry (ABC method) for expression of CD11b/c (microglial activation marker, OX42) and glial fibrillary acidic protein (astrocyte activation marker, GFAP). Study 2- Response thresholds on the von Frey test were recorded prior to and across a timecourse through 42 days after either avulsion, rhizotomy, or sham surgery (n=2 for sham and avulsion, n=4 for rhizotomy). Study 3- Response thresholds on the von Frey test were recorded prior to and across a timecourse after avulsion surgery. Beginning on Day 12 post-surgery, rats (n=4/group) received once daily s.c. injections of either the glial activation inhibitor AV411/Ibudilast (Sigma; 5-mg/kg or 10-mg/kg) or equivolume vehicle (corn oil; 0.5-ml/kg). Daily drug treatment and intermittent behavioral testing continued through 33 days after surgery. 171.22 Ipsilateral Paw AV411 vs Vehicle Treatment Study 3: The glial modulator AV411 reverses bilateral allodynia induced by unilateral T13/L1 dorsal root avulsion. Daily administration of AV411 reversed allodynia induced by avulsion. The reversal begins after 5-6 injections and continues to reverse behavior until day 33. The contralateral paw began to resolve at day 28 and the ipsilateral paw was still sensitive at day 33. There were not differences between the 5mg and 10mg dose of AV411. Site of injury relative to the region modulating pain. Our preliminary data show an increase in immunoreactivity for GFAP (astrocyte activation marker) over 3 weeks. Ipsilateral Paw GFAP Contralateral Paw AV411 vs Vehicle Treatment Contralateral Paw Site of injury Quantification of changes in microglial activation is ongoing, however the microglia appear to be activated in response to avulsion. OX-42 Sham Ipsilateral Dorsal Horn OX-42 Avulsion Ipsilateral Dorsal Horn Spinal cord region modulating hind paw nociception Ipsilateral Paw Study 2: Unilateral T13/L1 avulsion produces sustained mechanical allodynia beyond that induced by rhizotomy. Allodynia was reliably observed 14 days after avulsion and rhizotomy procedures, compared to sham controls. Interestingly, rats that received avulsion surgery had absolute thresholds consistently below 1 gram whereas rats that received rhizotomy surgery had absolute thresholds consistently above 1 gram. Avulsion rats were still sensitive 42 days post-surgery while rhizotomy rats were close to their baseline levels. 18 days post unilateral T13/L1 avulsion surgery. Rats receiving avulsion surgery recover quickly. We allow them to recover from surgery in a padded box so that they do not injure their backs upon waking up. Both shams and avulsion animals often have some motor difficulty up to day 7 post-surgery and by day 14 both sham and avulsion animals show no motor deficits. They are always curious about their surroundings and do not protest being handled immediately after surgery. 21 days post unilateral T13/L1 avulsion surgery. Rats receiving avulsion surgery exhibit normal rat behavior. They are not aggressive, and they eat, drink, and groom normally. They are able to walk around on hard surfaces and show only very minor motor defects. Contralateral Paw Avulsed dorsal roots.. After performing a laminectomy, the dorsal root nerve is avulsed. The small red dots on the spinal cord represent where the dorsal roots were avulsed. Shows where the rootlets were avulsed Study 1: Unilateral T13/L1 avulsion produces sustained mechanical allodynia and may activate glia. Unilateral T13/L1 Avulsion induced reliable mechanical allodynia by day 14. Ipsilateral allodynia was maintained through the remainder of the 30 day timecourse. Mild contralateral allodynia was reliably observed on day 21, and remained through the end of the experiment. We are currently processing spinal cord tissue for immunohistochemistry in order to assess changes in glial activation. Facial allodynia at 24 hours • In sum… • We have developed a model of spinal cord injury involving T13/L1 avulsion. This procedure induces a progressively developing mechanical allodynia that lasts at least 1.5 months after surgery. This procedure is clinically relevant given that human avulsion injuries are associated with a high rate of chronic pain. The advantages of the model are numerous, including: (a) avoiding behavioral confounds that necessarily occur with paralysis associated with prior spinal cord injury models, and (b) lack of urinary retention and associated health issues. • Mechanical allodynia (Bigbee et al., Exp. Neurol., 2007) has previously been observed in ventral root avulsion models, but this is the first time a unilateral T13/L1 dorsal root avulsion has been used to model exaggerated pain associated with spinal cord injury. We show here that the avulsion of T13 and L1 dorsal rootlets induces bilateral mechanical allodynia in the hind paws lasting upwards of 1.5 months. This robust allodynia is sustained beyond that induced by rhizotomy. • We show that injury at the T13 and L1 dorsal root entry zones results in exaggerated pain in the hind paws, which are innervated primarily by L5 and L6. That is, we are showing pain below the level of injury. We are currently developing methods by which to assess exaggerated pain at levels between the injury and hind paws. • Consistent with peripheral nerve injury models and other spinal cord injury models, our initial observation is that our avulsion surgery activates glia. Classically, astrocytes gradually become activated and sustain that level of activation over time, thereby maintaining the exaggerated pain state. Astrocyte activation appears to follow this pattern in our avulsion model. Characterization of the microglial activation profile in our avulsion model is in progress. • This is the first time AV411 has been shown to reverse mechanical allodynia in an animal model of spinal cord injury. The reversal of allodynia by AV411 strongly points to the involvement of glial cells in the induction and maintenance of exaggerated pain. Future studies will elucidate these mechanisms in hopes that AV411 could prove to be a novel treatment option for patients suffering from exaggerated pain associated with spinal cord injury. • Acknowledgements • We would like to thank Ken Lynch and Nicole Crysdale for their technical assistance. This work is supported by Craig Center for Spinal Cord Injury Research and NIH grants DA01542 & DA022042. • Contact Information: ellisal@colorado.edu IL1 TNF IL1