
26年度環境応答生理学第3回 (5 / 21) 塩ストレス環境とイオン輸送系 その2 (イオン輸送の最新研究成果) 、 環境応答生理におけるシグナル伝達、細胞死
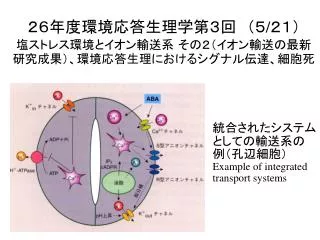
26年度環境応答生理学第3回 (5 / 21) 塩ストレス環境とイオン輸送系 その2 (イオン輸送の最新研究成果) 、 環境応答生理におけるシグナル伝達、細胞死. 統合されたシステムとしての輸送系の例(孔辺細胞) Example of integrated transport systems. カリウム輸送系とナトリウムの吸収. 植物(シロイヌイナズナ)のカリウム輸送系遺伝子. オオムギ切断根のカリウム吸収速度と培地カリウム濃度の関係 複数の輸送体の寄与を示唆 Epstain, Nature 212:1324 (1966). 塩ストレスでは・・・. Na +. 代謝と

26年度環境応答生理学第3回 (5 / 21) 塩ストレス環境とイオン輸送系 その2 (イオン輸送の最新研究成果) 、 環境応答生理におけるシグナル伝達、細胞死
E N D
Presentation Transcript
26年度環境応答生理学第3回 (5/21)塩ストレス環境とイオン輸送系 その2(イオン輸送の最新研究成果)、環境応答生理におけるシグナル伝達、細胞死 統合されたシステムとしての輸送系の例(孔辺細胞) Example of integrated transport systems
カリウム輸送系とナトリウムの吸収 植物(シロイヌイナズナ)のカリウム輸送系遺伝子 オオムギ切断根のカリウム吸収速度と培地カリウム濃度の関係 複数の輸送体の寄与を示唆 Epstain, Nature 212:1324 (1966) 塩ストレスでは・・・ Na+ 代謝と 膜電位の 攪乱 Na+↑ K+ K+↓
代替(有益) カリウム不足、適度なナトリウムは・・・ • カリウムを代替して生育改善 • (「植物栄養学」第2版) • イネではOsHKT2;2がこのNa透過を担っているらしい • Plant Physiology 152:341 (2010)
out in K+ Na+ 課題論文 Sience 270:1660 では・・・ out in K+ X Na+ コムギHKT (Na/K 共輸送) Na+透過に関与するのは A240とL247
Plant Physiology152:341, 2010 イネHKTは?OsHKT2;1 : K非依存Na輸送 OsHKT2;2: K依存的Na輸送 Na依存的K輸送 (多量のNaがあると、もっぱらNaを輸送) シロイヌナズナHKTもっぱらNaのみを輸送 その役割は? (タバコ培養細胞に発現させて測定) Plant Physiology136:2457, 2004 K欠乏下でOsHKT2;1発現→Kの代替をするNa輸送(EMBOJ 26:3003, 2007)
Schematic summaries of Na+ influx pathways into saline roots Primary protective mechanisms mediated by Na+ transporters on important biological membranes Rice誌5:11 (2012)) 注意:植物種によって、イオン選択性は異なる Apoplastic bypath flow
環境応答生理におけるシグナル伝達 環境 植物ホルモン レセプター シグナル伝達 レセプター 転写制御 (translational regulation) 遺伝子発現 タンパク質 代謝レベル 転写後制御 (post-translational regulation) 表現型
植物ホルモンエチレン • 果実の成熟制御 • 成長(方向、時期)制御 エチレン受容体(ETR1) 二成分(ヒスチジンカイネース)系 エチレン応答: • フック • 太く短い茎 • 太く短い主根 MAPKカスケード相当シグナル伝達 (後述) かなり簡略化している (実はもっと複雑) Two-Component Signal Transduction Pathways in Arabidopsis. Hwang et al. Plant Physiology, 2002, 129:500–515
シロイヌナズナの例 ここにMAPKシグナル経路 (後述) Figure 9. Model of the two-component signal transduction pathways in Arabidopsis. The cytokinin signal is perceived by multiple His protein kinases at the plasma membrane. Upon perception of the cytokinin signal, His protein kinases initiate a signaling cascade via the phosphorelay that results in the nuclear translocation of AHPs (Hwang and Sheen, 2001). Activated AHPs may interact with sequestered ARRs or ARR complexes, transfer the phosphate to the receiver domain of its cognate B-type ARR, releasing these activation-type ARRs from putative repressors in the nucleus. The dephosphorylated AHP shuttles back to the cytosol, where it can be rephosphorylated. The liberated ARRs bind to multiple cis elements in the promoter of target genes. The activation of the repressor-type ARRs as primary cytokinin response genes provides a negative feedback mechanism. In addition to the CTR1 signaling pathway, additional ethylene signaling pathways could be mediated by two-component components (Lohrmann and Harter, 2002). Red light and cytokinin signaling is converged at ARR4. ARR4 stabilizes the active form of PHYB by inhibiting dark reversion (Sweere et al., 2001). Stress and Glc may also modulate two-component signaling (Urao et al., 1998; F. Rolland and J. Sheen, unpublished data). RD, Response domain; BD, DNA binding domain; AD, transactivation domain; PM, plasma membrane; N, nucleus; R, putative repressor; FR, far-red light
低温ストレス応答の場合 らん藻で研究が進んでいる、ヒスチジンカイネース系 Hypothetical scheme for the pathway for low-temperature signal transduction in SynechocystisSuzuki et al. Biochem. Soc. Trans. (2000) 28, 628–630 どうやって証明? ↓ HiK33欠損変異体を作成、解析
らん藻での高浸透圧ストレス応答とヒスチジンカイネース系らん藻での高浸透圧ストレス応答とヒスチジンカイネース系 Five Histidine Kinases Perceive Osmotic Stress and Regulate Distinct Sets of Genes in Synechocystis Paithoonrangsarid et al. J. Biol. Chem., (2004) 279: 53078-53086
植物の MAPKinase系 Nature 451, 789-795 (2008) "Dual control of nuclear EIN3 by bifurcate MAPK cascades in C2H4 signalling." Yoo et al. なぜカスケードか? Model of the bifurcate MAPK cascades in ethylene signalling. The two EIN3 phosphorylation sites (P) with opposite functions are marked. Without ethylene, CTR1 directly or indirectly inactivates MKK9–MPK3/6 and probably activates downstream MAPKs to phosphorylate T592 to promote EIN3 degradation. Ethylene inactivates CTR1 for MKK9–MPK3/6 activation and T174 phosphorylation to stabilize EIN3. Arrow and blunt ends indicate positive and negative regulations, respectively. ACS, ACC synthase; MKKK, MAPKKK; C, cytoplasm; N, nucleus. (名古屋大学植物発生学研究室HPより) 増幅 調節 分岐・統合
アブシジン酸 ABAとDREB: 低温と乾燥に対するシロイヌナズナの応答 ABAは農業生産上の重要な形質(種子(形成と発芽制御)、ストレス耐性)に関与 受容 → 伝達 転写因子 → シス因子 →
アブシジン酸受容体 <2006~7年にかけての発表(natureとscience)は、いずれも撤回された。> 2009年に報告されたものが本物 • RCAR/PYR/PYLタンパク質(タイプ2C protein phosphatase 結合蛋白) Science 324:1064 および 同 324:1068 (2009) • RCAR1: Regulatory Component of ABA receptor1 (酵母TwoHybrid) PYR1: PYRABACTIN RESISTANCE 1 (ABAアゴニスト:活性化物質) S. Y. Park, P. Fung, N. Nishimura, D. R. Jensen, H. Fujiiet al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. 発芽成長阻害剤 タンパク質脱リン酸化酵素PP2C = ABI その変異体(abi)がABA Insensitive(ABA関連反応が失われる) として知られていた 他のABA関連変異体aba:ABA合成酵素の変異 http://www.sciencemag.org/content/324/5930/1068.full
A model of (+)-ABA-induced ABI1 regulation by PYL1. K Miyazonoet al. Nature462, 609-614 (2009) doi:10.1038/nature08583 ABAの生理作用1:種子休眠 ABAの生理作用2:ストレス耐性 10個ある (シロイヌナズナ) ABAとトランス因子、シス因子 (既出図の一部) ストレス耐性関連遺伝子の発現 ABAの生理作用3:気孔閉鎖 OST1=SnRK2.6=SnRK2E
Phenotypes of srk2d/i and srk2d/e/i mutants. Nakashima K et al. Plant Cell Physiol 2009;50:1345-1363 SnRK2D = SnRK2.2 SnRK2E = SnRK2.6 SnRK2I = SnRK2.3 ABA 種子 休眠 三重変異体 種子 休眠解除 十重変異! Arabidopsis decuple mutant reveals the importance of SnRK2 kinases in osmotic stress responses in vivo Fujii H et al. PNAS 2011;108:1717-1722
(引き続き) Arabidopsis decuple mutant reveals the importance of SnRK2 kinases in osmotic stress responses in vivo 236またはt :snrk2.2/3/63重変異体 (2.2と2.3と 2.6の3つが欠損しているという意味) quad:snrk2.1/4/5/10の4重変異体 quin: snrk2.1/4/5/9/10の5重変異体 sまたはsep:snrk2.1/4/5/7/8/9/10 の7重変異体 dまたはdec:10重変異体(すべて欠損) 2W:2:2だけが機能していて他の9つは欠損変異体 WT:野生型(すべてのSnRKが機能している) Seedling growth under osmotic stress. さらなる役割分担/微調節 Proline: 浸透圧ストレスに応答して蓄積 ABA 浸透圧ストレス 耐性 十重変異体 浸透圧ストレス 耐性
ホルモン制御系やシグナル伝達系はなぜ複雑なのか?ホルモン制御系やシグナル伝達系はなぜ複雑なのか? 多重性(因子、経路) クロストーク 「植物ホルモンのシグナル伝達」より Plant Physiology 154:567 (2010) BMC Plant Biology 11:163 (2011) 現代の自動車 + AT ABS キックダウン制御 ・・・・・・ 初期の自動車 アクセル ブレーキ
ストレス応答および細胞死とその分子機構 核 自爆装置 “オフ” 病理的刺激 (ストレス) 組織の恒常性の維持 生理的細胞死 シグナル 有害細胞の除去 (感染時のHR) 核 自爆装置 “オン” 形態形成 疾患(障害) 「科学」68:223(1998) 「細胞の持つ自殺機構 アポトーシスを制御する」 (辻本)を改変 プログラム細胞死(PCD)
↑ もともとの動物細胞での、形態的特徴に基づいたアポトーシス(特徴的な細胞死)の定義(animal cells) 植物でプログラム細胞死が起こる場所/機会 (Tissue and timing in plants) • プログラム細胞死(PCD)発生プログラムに従った、遺伝子や酵素の発現・活性調節を伴った生理的細胞死。シグナル伝達系が関与。 • 植物の場合核とDNA、膜の構造変化はおこるが、動物のアポトーシスに特有な形態変化の多くは見られない。 PCDにつながる細胞内シグナル伝達がある。機能は、かなり動物と共通。遺伝子は、大部分が異なる。
オオムギ根での塩ストレス誘導性細胞死 Salt stress-induced cell death高濃度の塩ストレスによる根の伸長成長の停止は細胞核の崩壊を伴う細胞死を引き起こしていた PCD in normal condition • 道管の形成 (xylem) • 破生間隙の形成(aerenchyma) • Control (up) • 500mM NaCl 12 hr (middle) • 500mM NaCl 24 hr (down)
中程度の塩ストレスによる根の伸長成長の停止中程度の塩ストレスによる根の伸長成長の停止 • 細胞は死んでいない • 膨圧の減少による伸長する力の消失 • 伸長域での形態異常→細胞壁の問題? • 細胞壁微小管は正常に存在するらしい
強い塩ストレスを受けた根の細胞死 Nuclear DNA degradation
TUNELTdT-mediated dUTP nick end labelingTdT: terminal deoxynucleotidyl transferase
DAPI TUNEL Control Stress 1h
塩ストレス誘導性細胞死は単細胞でも起こる Salt causes ion disequilibrium-induced programmed celldeath in yeast and plants Plant Journal (2002) 29: 649 その他に・・・ • TUNEL反応も見られる • 塩ストレス応答シグナル伝達系(CNB)が欠損すると、耐塩性が弱くなり、塩ストレス誘導性PCDが増える • 動物由来のBcl-2は、塩ストレス誘導性PCDを抑制できる
VPEというproteinaseが重要らしい(VPE: Vacuolar Processing Enzyme) N. benthamiana plants were infected with TMV on halves of their leaves (indicated by asterisks). (Hatsugai et al. Science 305:855, 2004のFig.2より) Non-silenced (pPVX) ↓ ← TMV感染HR(TNV封じ込め) VPE-silenced (pPVX: VPE) ↓ ← TMV感染HRおこらず(→全身感染へ) A model of “point-of-no-return” 京都大学大学院理学研究科 西村いくこ 研究室 http://www.jst.go.jp/kisoken/seika/zensen/08nishimura/index.html
細菌の感染後に細胞内の液胞の膜と細胞膜とが融合し、細胞の中と外がつながるトンネルが形成される細菌の感染後に細胞内の液胞の膜と細胞膜とが融合し、細胞の中と外がつながるトンネルが形成される 細菌感染に応答して誘導される膜融合(トンネル形成)とそれによる抗菌物質の放出と過敏感細胞死のモデル 京都大学大学院理学研究科 西村いくこ 研究室 http://www.kyoto-u.ac.jp/ja/news_data/h/h1/news6/2009/091015_1.htm 植物が本来もつ自己防衛能力を強化させる技術 細胞死の研究 ストレス誘導性細胞死の抑制/制御技術
動物細胞由来の細胞死誘導遺伝子 動物ホルモンDEX応答性のプロモーターの支配下にBax遺伝子等を入れて形質転換 Figure 9. Scheme of the Role of the BI-1 Protein in the ROS-InducedCell Death Pathway. ROS cause mitochondrial permeability transition, ATP depletion, releaseof cytochrome c from the mitochondrial membranes, and lipid peroxidation,which cause ion leakage and cell death. BI-1 proteins,as components of the ER membrane, are involved in counteracting “oxidative stress.” Figure 2. Suppression of Ion Leakage from Detached Leaves of Transgenic Arabidopsis Expressing Both Bax and AtBI-1. Dissection of Arabidopsis Bax Inhibitor-1 Suppressing Bax–, Hydrogen Peroxide–, and Salicylic Acid–Induced Cell Death. Kawai-Yamada et al. The Plant Cell, 16:21–32, 2004
PCDの分子機構: 動物細胞の場合 Death Domainを持つレセプター Death Effector Domain を持つ Caspase Downstream Caspase Nature Reviews Molecular Cell Biology 4, 552-565 (2003)