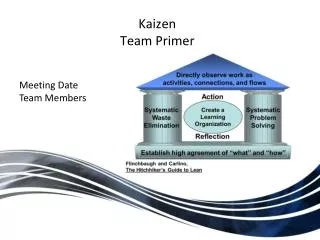
Primer Design: Size
Primer Design: Size. Genome Sizes: Prokaryote genome size: M genitalium = 0.5 million bp; E coli = 4 million bp Eukaryote genome size: Yeast = 12 million ; human = 3 billion ; maize = 4 billion bp Primer complexity / size:


Primer Design: Size
E N D
Presentation Transcript
Primer Design: Size Genome Sizes: • Prokaryote genome size: • M genitalium = 0.5 million bp; E coli = 4 million bp • Eukaryote genome size: • Yeast = 12 million; human = 3 billion; maize = 4 billion bp Primer complexity / size: • 17-base oligo has “complexity” or number of possible sequences of 417 = 17 179 800 000=fold • 1/4000 chance of occurring randomly in E coli genome • 1/4 chance of occurring randomly in maize genome MAKE PRIMERS LONG ENOUGH FOR TARGET GENOMES
Primer Design: Sequence 1 Rules for Primer Design: (adapted from Innis and Gelfand, 1991): • primers should be 17-28 bases in length; • base composition should be 50-60% (G+C); • primers should end (3') in a G or C, or CG or GC: this prevents "breathing" of ends and increases efficiency of priming; • Tms between 55-70oC are preferred; • runs of three or more Cs or Gs at the 3'-ends of primers may promote mispriming at G or C-rich sequences (because of stability of annealing), and should be avoided; • primer self-complementarity (ability to form 2o structures such as hairpins) should be avoided. • 3'-ends of primers should not be complementary (ie. base pair), as otherwise primer dimers will be synthesised preferentially • Primers should not be too degenerate
Primer Design: Sequence 2 Genome Sequence Known:
Primer Design: Sequence 2 Primer Properties:
Primer Design: Degeneracy One makes degenerate primers – or effective mixtures of many primers of similar but non-identical sequence – to amplify DNA that is evolutionarily related to known sequence(s). Eg: for “evolutionary” PCR, or discovery / characterisation of genes in species related to a model organism; For detection / discovery of novel organisms, for example, in environmental investigations such as oceanic / hot spring water bacterial sampling, biofilm diversity etc. Method: multiply align as many related sequences as possible, choose conserved areas of sequence, design primers so as to maximise the probability of amplifying as many sequence relatives as possible Examples: Detection of related viruses in faecal samples; detection of new bacterial species by rDNA gene PCR
Detection Methods: • Ethidium bromide-stained agarose gel of PCR products from various grasses infected with mastreviruses • Southern blot of agarose gel probed with MSV-specific labelled DNA • Note different intensities of bands in (a) and (b): this is due to the sequences being more or less related to the probe sequence
Annealing 1: temperatures Usually set at ~5oC below Tm • Tm Determined by length and GC content of oligo Formula for Tm calculation: Melting Temperature Tm(oK)={ΔH/ ΔS + R ln(C)}, Or Melting Temperature Tm(oC) = {ΔH/ ΔS + R ln(C)} - 273.15 where ΔH (kcal/mole) : H is the Enthalpy. Enthalpy is the amount of heat energy possessed by substances. ΔH is the change in Enthalpy. In the above formula the ΔH is obtained by adding up all the di-nucleotide pairs enthalpy values of each nearest neighbor base pair. ΔS (kcal/mole) : S is the amount of disorder a system exhibits is called entropy. ΔS is change in Entropy. Here it is obtained by adding up all the di-nucleotide pairs entropy values of each nearest neighbor base pair. An additional salt correction is added as the Nearest Neighbor parameters were obtained from DNA melting studies conducted in 1M Na+ buffer and this is the default condition used for all calculations. ΔS (salt correction) = ΔS (1M NaCl )+ 0.368 x N x ln([Na+]) WhereN is the number of nucleotide pairs in the primer ( primer length -1). [Na+] is salt equivalent in mM. [Na+] calculation: [Na+] = Monovalent ion concentration +4 x free Mg2+. http://www.premierbiosoft.com/tech_notes/PCR_Primer_Design.html
Annealing 2: temperatures Basic Tm Calculations The simplest formula is as follows: Tm = 4°C x (G +C) + 2°C x (A + T) This formula is valid for oligos <14 bases and assumes that the reaction is carried out in the presence of 50mM monovalentcations. For longer oligos, the formula below is used: Tm = 64.9°C + 41°C x (G + C – 16.4)/N Where N is the length of the primer. http://www.promega.com/biomath/calc11.htm
Annealing 3: time and temperatures Actual temperature used best determined EMPIRICALLY TIME of annealing not important UNLESS primers >50 bases – the process completes in seconds
Annealing 4: time and temperatures Actual temperature used best determined EMPIRICALLY Eg: vary annealing temperature across heating block 35 – 50oCgradient TIME of annealing not important UNLESS primers >50 bases – the process completes in seconds
Nature & Length of Template Template molecule could be: • Chromosomal dsDNA(bacterial or eukaryote or viral) • Plasmid / viral cccDNA (ss or ds) • Single-stranded RNA(viral or cellular) • Double-stranded RNA(viral) Template could be: • Short: 100 – 500 bases (detection purposes) • Medium length: 1000 – 5000 bases (whole gene / whole viral genome amplification) • Long: 10 000 – 40 000 bases (mapping cell genomes / whole viral genome amplifications)
Nature & Length of Template 2: Effect on denaturation temperature: • Need to denature hot enough for long enough • Short templates denature quicker than long • DNA:DNA < DNA:RNA < RNA:RNA in melting temperature • Tm depends upon G+C content • Usable denaturation temperatures are 88 - 95oC Time: • DNA denatures in just a few seconds at the “strand separation temperature” – BUT as there may be a lag time due to tube material insulating the reactants, so: • around 30 sec at chosen temperature (88 – 95oC) is required • T1/2 of Taq pol is: • >2 hrs at 92.5oC • 40 minutes at 95oC • 5 minutes at 97.5oC
Elongation Time Elongation time (assuming temperature = 70oC) depends on: • LENGTH of target sequence: • Taq pol makes 2 kb in 60 sec • STAGE of reaction: • As reactants get reduced with high template concentration, longer times are required • As Taq pol activity falls, may need more time
Reaction Buffer 1 • This is almost always provided as a concentrated kit component (to dilute 1/10). It generally contains: • 10 – 50 mM Tris-Cl pH 8.3: NB: reverse transcriptase buffer works well also • <50 mM KCl • ~1.5 mM MgCl2 • *Gelatin or BSA to 100 μM • *Non-ionic detergent 0.01 – 0.1 % v/v * = or other dispersant / protein stabiliser
Reaction Buffer 2 Mg2+ concentration should be 0.5 – 2.5 mM > [dNTP] as otherwise is all chelated TITRATE Mg2+!!!
Reaction Buffer 3 • HELIX DESTABILISERS may be needed for HIGH G+C TEMPLATES: • Urea, formamide, dimethylformamide (DMF) or dimethylsulphoxide (DMSO) or glycerol all help; DMSO IS BEST • Additives also help for LONG TEMPLATES • Polyethylene glycol (PEG) can help when [template] is LOW.
How Many Cycles is Enough? • 30 cycles = 109-fold amplification • 40 cycles = 1012-fold amplification • 50 cycles – 1015-fold amplification….?? Innis & Gelfand, 1990 • 50 copies of target DNA: 40 – 45 cycles • 5 x 105 copies of target: 25 – 30 cycles
Plateau Effect in DNA Amplification: Theoretical 40 Actual 30 Log2[product] 20 10 50 40 30 20 10 Cycle number
How to Amplify More DNA? • Re-amplify reaction product in separate reaction with same primers (just add more reagents): • Advantage: further amplification, no reactant depletion • Disadvantage: further amplification of non-specific products • Re-amplify reaction product with DIFFERENT primers (nested primer PCR) after clean-up: • Advantage: further amplification WITHOUT same non-specific products • Disadvantage: more manipulation
Nested Primer PCR: 1. 30 cycle PCR with Primer Set 1: ~109-fold amplification 2. 30 cycle PCR with Primer Set 2: ~109-fold amplification Final theoretical amplication: ~1018-fold Eg: 1 molecule of 1 kb -> 1 ng (30 cycles) -> …1 gram??
How much can you amplify? • Theoretically: Consider 100ul reaction with 1uM each primer, 200uM each dNTP, 1000 bp target. Total possible conversion limited by [dNTP] = 0.8 uM assuming A=G=C=T = 51.2 ug [(0.8 x 640 000 x 10-6)/10 000] • Practically: only a few micrograms as this reaction cannot go to completion as [reactants] primer + dNTP DNA
Amount amplified depends upon [template]: • [Product] saturates for [template] above certain limits ONLY RE-AMPLIFY SMALL ALIQUOT OF 1st REACTION!! C. Soin‚, S. K. Watson, E. P. Rybicki, B. Lucio, R. M. Nordgren, C. R. Parrish, and K. A. Schat. Determination of the detection limit of the polymerase chain reaction for chicken infectious anemia virus. Avian.Dis. 37 (2):467-476, 1993.