Download
1 / 51
550 likes | 752 Views
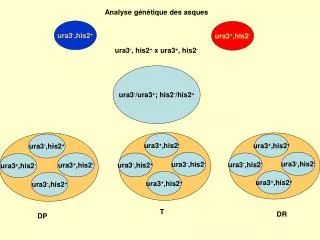
ura3 + ,his2 +. ura3 + ,his2 -. ura3 - ,his2 -. ura3 - ,his2 -. ura3 - ,his2 -. ura3 - ,his2 +. ura3 + ,his2 +. ura3 + ,his2 +. Analyse génétique des asques. ura3 - ,his2 +. ura3 + ,his2 -. ura3 - , his2 + x ura3 + , his2 -. ura3 - /ura3 + ; his2 - /his2 +. ura3 - ,his2 +.

E N D
ura3+,his2+ ura3+,his2- ura3-,his2- ura3-,his2- ura3-,his2- ura3-,his2+ ura3+,his2+ ura3+,his2+ Analyse génétique des asques ura3-,his2+ ura3+,his2- ura3-, his2+ x ura3+, his2- ura3-/ura3+; his2-/his2+ ura3-,his2+ ura3+,his2- ura3+,his2- ura3-,his2+ T DR DP
Comportements des gènes au cours de la méïose chez la levure S. cerevisiae : marqueurs physiquement indépendants
ura3-,his2+ ura3-, his2+ x ura3+, his2- ura3+,his2- his2+ his2+ ura3- ura3- his2+ his2+ his2- his2- ura3- ura3+ ura3+ his2- his2- ura3+ ura3+ ura3-,his2+ ura3+,his2- ura3+,his2- ura3+,his2+ ura3-,his2+ ura3-,his2- ura3-,his2- ura3+,his2+ DP ura3- DP = DR DR
ura3-,his2+ ura3-, his2+ x ura3+, his2- ura3+,his2- his2+ ura3- ura3- his2+ his2- his2- ura3+ ura3+ his2- his2- ura3+ ura3+ ura3+,his2- ura3-,his2- ura3-,his2+ ura3+,his2+ his2+ ura3- ura3- his2+ ura3-,his2+ Tetratypes ura3+,his2+ ura3+,his2- ura3-,his2- Les Tétratypes sont obtenus par 1 CO entre ura3 en cen, OU his2 et cen
Comportements des gènes au cours de la méïose chez la levure S. cerevisiae : marqueurs physiquement liés
his2+ ura3- his2- ura3+ ura3-,his2+ ura3+,his2- ura3+,his2- ura3-,his2+ DP
ura3+,his2+ ura3-,his2- ura3-,his2- ura3+,his2+ his2+ ura3- 2 CO entre ura3 et his2 affectant les 4 chromatides his2- ura3+ ura3-,his2+ ura3+,his2- ura3+,his2- ura3-,his2+ DR DP DP >>DR
ura3+,his2- ura3+,his2+ ura3-,his2- ura3-,his2- ura3-,his2- ura3-,his2+ ura3+,his2+ ura3+,his2+ his2+ ura3- 1 CO entre ura3 et his2 affectant 2 chromatides his2- ura3+ ura3-,his2+ ura3+,his2- ura3+,his2- ura3-,his2+ T DR DP DP >>DR
(Tx2 + DRx4) X 100 (DP + T + DR)x4 Fréquence de recombinaison entre ura3 et his2 F rec (ura3,his2) =
[arg+,leu-] [arg-,leu-] [arg-,leu+] [arg+,leu+] En résumé … [arg-,leu+] [arg+,leu-] [+] Le(les) mutation(s) récessives [arg+, leu+] [arg-, leu+] [arg-, leu+] [arg+, leu+] [arg-, leu-] [arg+,leu-] [arg+,leu-] [arg-, leu-] 2 spores [arg-], 2 spores [arg+] par asque : 1 couple d’allèle arg-/arg+ 2 spores [leu-], 2 spores [leu+] par asque : 1 couple d’allèle leu-/leu+
arg+,leu- arg-,leu- arg-,leu+ arg+,leu+ En résumé … [arg-,leu+] [arg+,leu-] ura3-, his2+ x ura3+, his2- [+] Le(les) mutation(s) récessives arg+, leu+ arg-, leu+ arg-, leu+ arg+, leu+ Arg-, leu- arg+,leu- arg+,leu- Arg-, leu- T DR DP
(Tx2 + DRx4) X 100 (DP + T + DR)x4 arg+,leu- arg-,leu- arg-,leu+ arg+,leu+ arg+, leu+ arg-, leu+ arg-, leu+ arg+, leu+ Arg-, leu- arg+,leu- arg+,leu- Arg-, leu- T DR DP • Si DP = DR : les gènes arg et leu sont génétiquement indépendants • Si DP >>> DR : les gènes arg et leu sont génétiquement liés Frec (arg,leu) =
Asques chez Neurospora crassa : cartographie du centromère Chez certains champignons, il y a une mitose à la suite de la méiose
Asques de Sordaria macrospora Spores noires x spores jaunes
A A A A a a A A a a a a A A a A A a a a Asque postréduit pour le gène étudié 1 CO entre A et le cen
23 asques analysables 10 asques préréduits
23 asques analysables 10 asques préréduits 13 asques postréduits
23 asques analysables 10 asques préréduits 13 asques postréduits Fpost = 100(13/23) = 56%
A A A A a a A A a a a a A A a A A a a a Fpost = 100(13/23) = 56% Dans un asque postréduit, 50% des spores sont recombinantes entre le gène et le centromère Frec(gène-cen) = Fpost(gène)/2 Frec(A-cen) = 28%
Distance à partir de laquelle un gène est indépendant du centromère? Asques postréduits Asques preréduits ????
Distance à partir de laquelle un gène est indépendant du centromère? Asques postréduits Asques preréduits La probabilité d’avoir l’un quelconque de ces 6 asques est identique (1/6) : la fréquence de postréduction est donc 100 x 4/6 = 66%. La fréquence de recombinaison de 33%. A partir de 33cM, un gène est génétiquement indépendant du centromère
En fonction de l’origine de la fleur sur la plante, la descendance est variable
Phénotype mosaïque lié à la présence d’ADN dans les chloroplastes , les mitochondries • Les parasites, intracellulaires peuvent aussi modifier les phénotypes (altération du sex ratio par exemple)
CHAP 4 Génétique des populations
Description génétique d’une population Une population est un ensemble d’individus d’une même espèce localisés dans une même région géographique. Dans une population locale, chaque individu peut trouver son partenaire sexuel. Comment se répartissent les différents allèles d’un gène au sein d’une population?
Le virus du Sida rentre dans les cellules en reconnaissant principalement un récepteur, le produit du gène CCR5. Cette protéine contient 352 aa. aa n° 171 K E G L H Y T C S S AAA GAA GGT CTT CAT TAC ACC TGC AGC TCT H F P Y S Q Y Q F W CAT TTT CCA TAC AGT GAG TAT CAA TTC TGG K N F Q T L K I V I AAG AAT TTC CAG ACA TTA AAG ATA GTC ATC L G L V L P L L V M TTG GGG GTG GTC CTG CCG CTG CTT GTC ATG V I C Y S G I L K T GTC ATC TGC TAC TCG GGA ATC CTA AAA ACT L L R C R N E K K R CTG CTT CGG TGT CGA AAT GAG AAG AAG AGG aa n° 230
L’allèle CCR5-Δ32 correspond à une délétion de 32 nucléotides K E G L H Y T C S S AAA GAA GGT CTT CAT TAC ACC TGC AGC TCT K E G L H Y T C S S H F P Y S Q Y Q F W CAT TTT CCA TAC AGT GAG TAT CAA TTC TGG H F P Y K N F Q T L K I V I AAG AAT TTC CAG ACA TTA AAG ATA GTC ATC I K D S H L G L V L P L L V M TTG GGG GTG GTC CTG CCG CTG CTT GTC ATG L G A G P A A A C H V I C Y S G I L K T GTC ATC TGC TAC TCG GGA ATC CTA AAA ACT G H L L L G N P K N L L R C R N E K K R CTG CTT CGG TGT CGA AAT GAG AAG AAG AGG S A S V S K stop CCR5 CCR5-Δ32 Décalage du cadre de lecture
La protéine CCR5-Δ32 est non fonctionnelle. • L’allèle CCR5-Δ32 a été identifié chez des individus séropositifs chez qui la maladie ne s’est pas déclarée pendant au moins 10 ans. 40% des personnes séropositives, non malade, sont hétérozygotes pour l’allèle CCR5-Δ32.
Etude de la répartition des allèles CCR5 et CCR5-Δ32 dans un échantillon de la population française CCR5/ CCR5 : 795 CCR5-Δ32/ CCR5 : 190 CCR5-Δ32/ CCR5-Δ32 015 Total : 1000 • Pourquoi y a t’il peu d’homozygotes CCR5-Δ32/ CCR5-Δ32?
CCR5/ CCR5 (A/A) : 795 CCR5-Δ32/ CCR5 (a/A) : 190 CCR5-Δ32/ CCR5-Δ32 (a/a): 015 Total : 1000 Fréquence des génotypes : Fréquence de A/A dans la population = 0,795 Fréquence des a/A dans la population = 0,19 Fréquence des a/a dans la population = 0,015 Fréquence des allèles dans la population : pour 1000 individus, • 795 A/A , soit 1590 allèles A • 190 A/a, soit 190 allèles A et 190 allèles a • 15 a/a, soit 30 allèles a Fréquence de l’allèle A = 100x(1590 + 190)/2000 = 0,89 Fréquence de l’allèle a = 100x(190 + 30)/2000 = 0,11
La fréquence des génotypes résulte-t’elle d’une répartition aléatoire des gamètes dans la population ? Fréquence de l’allèle A = 100x(1590 + 190)/2000 = 0,89 Fréquence de l’allèle a = 100x(190 + 30)/2000 = 0,11 a 0,11 A 0,89 A 0,89 A/a : 0,0979 A/A : 0,7921 a 0,11 A/a : 0,0979 a/a : 0,0121 Répartition aléatoire A/A = 0,7921 A/a = 0,1958 a/a = 0,0121 Fréquence observée A/A = 0,795 A/a = 0,190 a/a = 0,015 La répartition des allèles A et a dans la population française est donc le fait du hasard
Equilibre de Hardy et Weinberg Soit A et a les deux allèles possibles présents dans un locus La fréquence allélique de A est p; celle de a est q Pour la population étudiée, on a p + q = 1 (car ce sont les seuls deux allèles possibles) Quels seraient les fréquences de A et a à la génération suivante ?
La fréquence des allèles A et a chez le mâle ou la femelle est la même que dans la population. C’est donc aussi la fréquence des gamètes dans la population Mâle A (p) a (q) A (p) Femelle a (q) Génération suivante Génération actuelle
Mâle A (p) a (q) A (p) A/A (p2) A/a (pq) Femelle a (q) a/A (qp) a/a (q2) La fréquence des allèles A et a chez le mâle ou la femelle est la même que dans la population. C’est donc aussi la fréquence des gamètes dans la population A la génération suivante : • Fréquence des génotypes : • Fréquence du génotype A/A : p2 • Fréquence du génotype A/a : pq + qp = 2pq • Fréquence du génotype a/a : q2
Mâle A (p) a (q) A (p) A/A (p2) A/a (pq) Femelle a (q) a/A (qp) a/a (q2) La fréquence des allèles A et a chez le mâle ou la femelle est la même que dans la population. C’est donc aussi la fréquence des gamètes dans la population A la génération suivante : • Fréquence des génotypes : • Fréquence du génotype A/A : p2 • Fréquence du génotype A/a : pq + qp = 2pq • Fréquence du génotype a/a : q2 • Fréquence des allèles : • Fréquence de A : p2 + 2pq/2 = p(p+q) = p • Fréquence de a : q2 + 2pq/2 = q(p+q) = q
En résumé A la génération de départ : Fréquence de A = p Fréquence de a = q Les fréquences alléliques ne bougent pas. La structure génétique de la population reste identique A la génération suivante : Fréquence de A = p Fréquence de a = q Equilibre de Hardy et Weinberg 1908 Cette loi décrit une population qui n’évolue pas!!!
Comment varient les fréquences des génotypes selon celles des allèles, dans la situation décrite par Hardy Weinberg ?
Comment varient les fréquences des génotypes selon celles des allèles, dans la situation décrite par Hardy Weinberg ? Exemple f(A) = 0,9 f(a) = 0,1 f(A/A) = 0,92 = 0,81 f(A/a) = 2x0,9x0,1 = 0,18 f(a/a) = 0,12 = 0,01
Les allèles rares (CCR5-Δ32) d’une population se retrouvent donc essentiellement sous forme hétérozygote
Les facteurs de variation des populations Une population est dite polymorphe pour un caractère si au moins deux types morphologiques différents sont observables dans cette population : polymorphisme génétique. • La mutation • Dérive génétique • Sélection
Mutation Chez les métazoaires les mutations sont somatiques/germinales Mutations neutres = silencieuse (sans impact sur le phénotype) Les mutations peuvent être favorables ou délétères
Mutation Le taux de mutation, µ, varie en fonction des espèces
Mutation Soit p et q la fréquence de deux allèles A et a dans une population. Si le taux de mutation de A vers a est µ, à chaque génération, la fréquence de l’allèle A , p, dans la population diminue de µp, tandis que celle de a augmente de µp. Ainsi, à chaque génération, la valeur de p diminue. On montre qu’après n génération à partir du moment, t0, où la fréquence de A est de p0, la fréquence de A, pn est : pn = p0e-nμ Si A disparaît, on dit que la mutation (l’allèle) a est fixée p(a) = 1; A est éliminé p(A) = 0
Chez l’homme 1 génération = 30 ans. A étant fixé, il faudrait à peu près 200000 génération pour que la fréquence de l’allèle A dans la population ne soit plus que de 15%. Le taux de mutation d’un gène n’est pas, à lui seul, suffisant pour expliquer les variations de structure des populations
Dérive génétique La loi de Hardy-Weinberg est établie dans une population de grande taille au sein desquels les croisements sont aléatoires (panmixie). Or, • Les populations ne sont pas infinies. • Les croisements ne se font pas toujours de manière aléatoire... Cela entraîne des distorsions vis à vis de l’équilibre de Hardy-Weinberg source de « dérive génétique aléatoire»
Exemples de dérive génétique : échantillonnage