
新教材课时分配
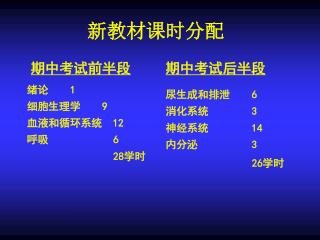
新教材课时分配. 期中考试前半段. 期中考试后半段. 绪论 1 细胞生理学 9 血液和循环系统 12 呼吸 6 28 学时. 尿生成和排泄 6 消化系统 3 神经系统 14 内分泌 3 26 学时. 第三章 细胞间信息传递. 细胞间信息传递的主要形式. 第一节 间隙连接与电传递. 1. 间隙连接结构、通透性和功能.

新教材课时分配
E N D
Presentation Transcript
新教材课时分配 期中考试前半段 期中考试后半段 绪论 1细胞生理学 9血液和循环系统 12呼吸 6 28学时 尿生成和排泄 6消化系统 3神经系统 14内分泌 3 26学时
第三章 细胞间信息传递 细胞间信息传递的主要形式
第一节 间隙连接与电传递 1. 间隙连接结构、通透性和功能
间隙连接广泛分布于脊椎和无脊椎动物几乎所有类型的细胞中,尤其是需要进行快速通讯的细胞(如神经细胞和肌细胞),是构成电突触的结构基础,参与多种生理功能。间隙连接广泛分布于脊椎和无脊椎动物几乎所有类型的细胞中,尤其是需要进行快速通讯的细胞(如神经细胞和肌细胞),是构成电突触的结构基础,参与多种生理功能。 间隙连接作为低电阻的通道,可将众多细胞在电学上联系起来,使细胞电活动协调一致,参与细胞的兴奋、收缩和分泌等功能。 间隙连接还参与细胞分化、生长与发育的过程。胚胎发育早期,间隙连接能够协调细胞间的发育过程,诱导细胞向一定方向分化。不同发育阶段的细胞表达不同的间隙连接蛋白,作用也不相同。
电传递 • 双向传递 • 速度快 • 不易受影响
第二节 化学传递的一般规律 一、化学传递研究历史 最早证明化学传递存在的实验是“迷走物质”的发现. 1905年,剑桥大学生理学家Elliott提出有化学物质参与交感的兴奋传递,未被接受。 1921年奥地利生理学 Loewi家用实验证明“迷走物质”的存在。 在Dale的建议下用胆碱脂酶抑制剂延长“迷走物质”作用,证实 为乙酰胆碱 二人获1936年诺贝尔奖
二、化学传递的基本机制 (1)化学突触的结构 和分类 • 结构: • 突触前膜 • 突触间隙 • 突触后膜
大脑皮层突触的电镜照片 (引自Bern, Physiology, 2005) S1和S2分别是两个神经突触,Den代表树突。
(1)A 轴突-轴突型突触 B 轴突-胞体型突触 C 轴突-树突型突触 (2)四种组合形式突触模式图 突触分类:
非定向、开放式突触传递 非突触性化学传递 (non-synaptic chemical transmission 曲张体小泡内递质与效应细胞间的特殊联系。 • 没有经典的突触结构; • 不存在一对一的支配关系; • 递质弥散距离大,传递时间长; • 作用部位发散,无特定的靶点; • 效应器能否发生作用取决与有无 相应的受体
(2)突触传递的原理 基本同神经-肌接头的传递过程。 突触后膜上产生的电位称为 突触后电位(postsynaptic potential).
兴奋性突触后电位 • (excitatory postsynaptic potential EPSP)
特点:电位大小取决于传入神经刺激强度的大小特点:电位大小取决于传入神经刺激强度的大小 产生过程: →突触前膜释放兴奋性递质 传入神经冲动到达末梢 →膜对Na+、 K+,尤其Na+ 递质与后膜特异受体结合 →膜电位降低,出现局部去极化 (EPSP) 的通透性增加 EPSP达一定程度,在轴突始段产生动作电位→ 动作电位沿神经传导
抑制性突触后电位 • (Inhibitory postsynaptic potential IPSP) • 所有的突触后抑制都是由抑制性中间神经原的活动引起的。 机制:通过抑制性突触后电位 (IPSP) 的形成 →神经末梢释放抑制性递质 抑制性神经元兴奋 递质与后膜特异受体结合 →膜对 K+ 、Cl- 或Cl- 的通透性增加 →膜电位超极化即IPSP 突触后膜兴奋性降低效应 产生抑制效应
突触后神经元的兴奋和抑制 兴奋性突触后电位( EPSP )和抑制性突触后电位( IPSP )
三、化学突触传递的信使分子——神经递质 1.神经递质的概念 一个化学物质被定为神经递质,必须具备五个条件: (1) 突触前神经元内具有合成递质的前体和酶系。 (2) 它储存于小泡内不被酶降解,神经冲动到达能释放。 (3) 其作用在后膜上,人为引入可引起相同的生理效应。 (4) 存在有使此物质失活的酶或其他环节。 (5) 有受体激动剂或受体的阻断剂能模拟剂或阻断作用。 2. 调质的概念: 3. 递质的共存: 戴尔原则(Dale’s principle) 递质共存
一氧化氮、一氧化碳等也在突触传递中发挥关键作用, 但它们并不严格符合经典神经递质标准。 目前发现的内源性气体信使分子(gaseous messenger)主要包括一氧化氮(NO)、一氧化碳(CO)和硫化氢(H2S)三种。
“NO NEWS IS GOOD NEWS ” Robert F. Furchgot Ferid Murad Louis J. Ignarro 20 世纪 80 年代初期,Robert F. Furchgott 发现血管内皮细胞通过释放一种被他命名为 “ 内皮细胞舒张因子 ” 维持血管张力和抑制血小板聚集。Furchgott 和 Louis J. Ignarro ( 1987 )分别独立地提出 EDRF 的本质是一氧化氮( nitric oxide, NO )1992 年 NO 被《 SCIENCE 》杂志评选为本年度明星分子,同期《 SCIENCE 》发表了一篇被冠以有趣标题 “ NO NEWS IS GOOD NEWS ” 的专论,以强调这一研究领域的重要性和新颖性。 1998 年 Furchgott 、 Ignarro 及 Ferid Murad 获得了诺贝尔生理学 或 医学奖。
2.神经递质的合成与释放 乙酰胆碱和氨基酸等小分子递质在电子密度低、直径40 ~ 60 nm的突触囊泡(synaptic vesicle)内贮存, 神经肽等大分子神经递质则在电子密度高、直径为90 ~ 250 nm的称为分泌颗粒的大囊泡中贮存。
3.神经递质的量子释放 除了视网膜感光细胞和一小部分释放脂溶性递质(如前列腺素、NO等)的细胞外,几乎所有神经元都遵循量子式释放的规律。这就是神经递质释放的“量子释放理论”。
四、受体 受体(receptor)是指靶细胞上能与神经递质或激素等化学信号分子(配体,ligand)结合并产生信号传递或转换作用的特异性蛋白质分子或复合体。
第三节 离子通道型受体介导的突触传递 一、N 型乙酰胆碱受体介导的突触传递
二、离子通道型谷氨酸受体介导的兴奋性突触传递二、离子通道型谷氨酸受体介导的兴奋性突触传递 谷氨酸受体:促代谢型谷氨酸受体;11种亚型 离子型谷氨酸受体: NMDA受体 NON- NMDA 受体:AMPA 和KA受体
AMPA受体 • 主要由GluR1、GluR2、GluR3、G1uR4 4种亚基通过不同的组合形成多种异构体。典型的受体中都包括一个GluR2亚基。 • AMPA受体介导谷氨酸快速兴奋性突触传递。AMPA受体激活后对Na+、K+通透性增加,使突触后神经元去极化。 • 在某些神经元,对钙高度通透。
Gly site Polyamine site Zn2+ site H+ MK-801 Mg2+
2.NMDA受体NMDA受体由NR1 和4 种不同的NR2 亚基(NR2A、NR2B、NR2C和NR2D)组成。 NMDA 受体对Ca2+ 的通透性大。钙调节一些离子通道的开启状态并能影响基因表达。过多的Ca2+ 进入细胞会引发细胞凋亡。激活NMDA受体能对突触后神经元产生持久而广泛的影响。现在认为,反复激活NMDA受体是神经系统高级功能如长时程记忆和学习的基础。
NMDA受体特征: 电压依赖的Mg2+ 阻断作用;对钙离子高度通透; 甘氨酸的协同剂作用; 多结合位点:激动剂、锌、H+, 磷 酸化、氧化还 原等
三、离子通道型谷氨酸受体介导的抑制性突触传递三、离子通道型谷氨酸受体介导的抑制性突触传递 1.离子通道型 -氨基丁酸受体 -氨基丁酸(GABA)是脊椎动物中枢神经系统最主要的抑制性神经递质之一。 根据药理学特性分为三类: -氨基丁酸(GABA) 受体: GABAA (促离子型受体):Cl-通道; Bicuculline敏感 GABAB (促代谢型受体):通过IP3、DG增加钾电导; GABAc (促离子型受体):Cl-通道,分布视觉通路; Bicuculline不敏感 三者都引起突触后膜超极化而产生抑制效应。
除了有GABA的结合位点外,还有与其他外源性镇静药物如苯二氮类药物和巴比妥类药物的结合位点。这些临床上广泛应用的镇静药物单独结合到受体上并没有显著效果,但与GABA同时结合则能显著增强GABAA 受体的抑制性作用。 苯二氮类药物可以增加Cl-通道开放的频率, 巴比妥类药物则能延长Cl- 通道开放的 时程。这样都会导致更多的Cl- 进入细胞,使神经元膜电位超极化
2.离子通道型甘氨酸受体 甘氨酸是一种最简单的氨基酸,是由脊髓和脑干中某些中间神经元释放的抑制性神经递质 甘氨酸受体:(GlyR) 由α 亚基、β 亚基构成的五聚体,与nAChR、GABAA 和5-羟色胺受体等具有很大的同源性,共同形成一个配体门控离子通道超家族。
低浓度的甘氨酸可以作为NMDA受体的共同配体,增强NMDA受体介导的反应;高浓度的甘氨酸,可以引起NMDA受体的内吞。低浓度的甘氨酸可以作为NMDA受体的共同配体,增强NMDA受体介导的反应;高浓度的甘氨酸,可以引起NMDA受体的内吞。
第四节 G蛋白耦联受体介导的突触传递 一、G 蛋白耦联受体及信号转导
二、肾上腺素受体的信号转导 根据受体的药理学反应不同,将肾上腺素受体分为和β 二种受体,介导不同的组织反映。 受体: 1 受体:主要分布于血管、瞳孔开大肌、胃肠及膀胱括约肌、立毛肌、泌尿生殖器的平滑肌和肝等 2 受体:主要分布于去甲肾上腺素神经末梢突触前膜以及血管等处的突触后膜
受体 1受体:主要分布在心脏以及肾的球旁细胞中 2受体:分布在血管平滑肌、细支气管平滑肌、膀胱和 子宫平滑肌、骨骼肌和肝中 3受体:主要分布在脂肪组织中。
三、M 型乙酰胆碱受体与信号传递 利用分子克隆技术已经鉴定出5种不同的M受体基因,分别编码 M1~M5 受体
M1、M4、M5受体:分布于脑内,调节某些认知或运动功能;M1、M4、M5受体:分布于脑内,调节某些认知或运动功能; M2 受体:分布于心脏窦房结和房室结、心房肌和心室肌细胞 M3 受体: 分布于胃肠道、输尿管的平滑肌和汗腺等;
有的组织中同时有两种或两种以上M受体,但却起着不同的生理作用。例如,心肌细胞M2受体与心肌收缩有关,而M1受体则可导致心率下降。有的组织中同时有两种或两种以上M受体,但却起着不同的生理作用。例如,心肌细胞M2受体与心肌收缩有关,而M1受体则可导致心率下降。
四、其他G 蛋白耦联受体介导的神经递质信号传递
1.代谢型谷氨酸受体(mGluR) 2.B型γ-氨基丁酸受体(GABAB受体) 3.多巴胺受体 受体亚型: D1 样 (D1 D5), 受体激活后升高cAMP水平; D2 样 (D2, D3, D4),受体激活后降低cAMP水平; 位于中枢三个部分:黑质-纹状体;中脑边缘系统; 结节-漏斗部;