Download
1 / 1
10 likes | 129 Views
c-Met, an autism spectrum disorder-associated gene, regulates dendritic spine structure and dendritic arborization in hippocampal neuron cultures Quanzhen Hao, Chol Seung Lim, Randall Walikonis* Department of Physiology & Neurobiology, University of Connecticut, Storrs, CT 06269. 44.14.

E N D
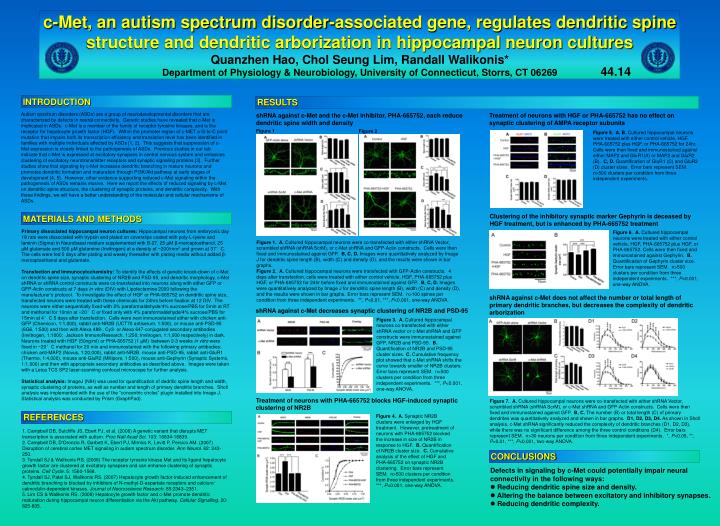
c-Met, an autism spectrum disorder-associated gene, regulates dendritic spine structure and dendritic arborization in hippocampal neuron cultures Quanzhen Hao, Chol Seung Lim, Randall Walikonis* Department of Physiology & Neurobiology, University of Connecticut, Storrs, CT 06269 44.14 INTRODUCTION RESULTS Autism spectrum disorders (ASDs) are a group of neurodevelopmental disorders that are characterized by defects in neural connectivity. Genetic studies have revealed that c-Met is implicated in ASDs. c-Met is a member of the family of receptor tyrosine kinases, and is the receptor for hepatocyte growth factor (HGF). Within the promoter region of c-MET a G-to-C point mutation that impairs both its transcription efficiency and translation level has been identified in families with multiple individuals affected by ASDs [1, 2]. This suggests that suppression of c-Met expression is closely linked to the pathogenesis of ASDs. Previous studies in our lab indicate that c-Met is expressed at excitatory synapses in central nervous system and enhances clustering of excitatory neurotransmitter receptors and synaptic signaling proteins [3]. Further studies show that signaling by c-Met increases dendritic branching in mature neurons and promotes dendritic formation and maturation through PI3K/Akt pathway at early stages of development [4, 5]. However, other evidence supporting reduced c-Met signaling within the pathogenesis of ASDs remains elusive. Here we report the effects of reduced signaling by c-Met on dendritic spine structure, the clustering of synaptic proteins, and dendritic complexity. With these findings, we will have a better understanding of the molecular and cellular mechanisms of ASDs. shRNA against c-Met and the c-Met inhibitor, PHA-665752, each reduce dendritic spine width and density Treatment of neurons with HGF or PHA-665752 has no effect on synaptic clustering of AMPA receptor subunits Figure 1 Figure 2 Figure 5. A, B. Cultured hippocampal neurons were treated with either control vehicle, HGF, PHA-665752 plus HGF, or PHA-665752 for 24hr. Cells were then fixed and immunostained against either MAP2 and GluR1(A) or MAP2 and GluR2 (B). C, D. Quantification of GluR1 (C) and GluR2 (D) cluster sizes. Error bars represent SEM. n>500 clusters per condition from three independent experiments. Clustering of the inhibitory synaptic marker Gephyrin is deceased by HGF treatment, but is enhanced by PHA-665752 treatment MATERIALS AND METHODS Primary dissociated hippocampal neuron cultures:Hippocampal neurons from embryonic day 18 rats were dissociated with trypsin and plated on coverslips coated with poly-L-lysine and laminin (Sigma) in Neurobasal medium supplemented with B-27, 25 μM β-mercaptoethanol, 25 μM glutamate and 500 μM glutamine (Invitrogen) at a density of ~200/mm2 and grown at 37°C. The cells were fed 5 days after plating and weekly thereafter with plating media without added β-mercaptoethanol and glutamate. Transfection and Immunocytochemistry:To identify the effects of genetic knock-down of c-Met on dendritic spine size, synaptic clustering of NR2B and PSD-95, and dendritic morphology, c-Met shRNA or shRNA control constructs were co-transfected into neurons along with either GFP or GFP-Actin constructs at 7 days in vitro (DIV) with Lipofectamine 2000 following the manufacturer’s protocol. To investigate the effect of HGF or PHA-665752 on dendritic spine size, transfected neurons were treated with these chemicals for 24hrs before fixation at 12 DIV. The neurons were either sequentially fixed with 2% paraformaldehyde/4% sucrose/PBS for 5min at RT and methonal for 10min at −20°C or fixed only with 4% paraformaldehyde/4% sucrose/PBS for 15min at 4°C 5 days after transfection. Cells were next immunostained either with chicken anti-GFP (Chemicon, 1:1,000), rabbit anti-NR2B (UCT70 antiserum, 1:500), or mouse anti-PSD-95 (6G6, 1:500) and then with Alexa 488-, Cy3- or Alexa 647-conjugated secondary antibodies (Invitrogen, 1:1000; Jackson ImmunoResearch, 1:250; Invitrogen, 1:1,000 respectively) in dark. Neurons treated with HGF (50ng/ml) or PHA-665752 (1 μM) between 2-3 weeks in vitro were fixed in −20°C methanol for 20 min and immunostained with the following primary antibodies: chicken anti-MAP2 (Novus, 1:20,000), rabbit anti-NR2B, mouse anti-PSD-95, rabbit anti-GluR1 (Thermo, 1:4,000), mouse anti-GluR2 (Millipore, 1:500), mouse anti-Gephyrin (Synaptic Systems, 1:1,000) and then with appropriate secondary antibodies as described above. Images were taken with a Leica TCS SP2 laser-scanning confocal microscope for further analysis. Statistical analysis: ImageJ (NIH) was used for quantification of dedritic spine length and width, synaptic clustering of proteins, as well as number and length of primary dendritic branches. Sholl analysis was implemented with the use of the “concentric circles” plugin installed into Image J. Statistical analysis was conducted by Prism (GraphPad). Figure 6. A. Cultured hippocampal neurons were treated with either control vehicle, HGF, PHA-665752 plus HGF, or PHA-665752. Cells were then fixed and immunostained against Gephyrin. B. Quantification of Gephyrin cluster size.Error bars represent SEM. n>500 clusters per condition from three independent experiments. ***, P<0.001, one-way ANOVA. Figure 1. A. Cultured hippocampal neurons were co-transfected with either shRNA Vector, scrambled shRNA (shRNA ScrM), or c-Met shRNA and GFP-Actin constructs. Cells were then fixed and immunostained against GFP. B, C, D. Images were quantitatively analyzed by Image J for dendritic spine length (B), width (C) and density (D), and the results were shown in bar graphs. Figure 2.A. Cultured hippocampal neurons were transfected with GFP-Actin constructs. 4 days after transfection, cells were treated with either control vehicle, HGF, PHA-665752 plus HGF, or PHA-665752 for 24hr before fixed and immunostained against GFP. B, C, D. Images were quantitatively analyzed by Image J for dendritic spine length (B), width (C) and density (D), and the results were shown in bar graphs.Error bars represent SEM. n>100 spines per condition from three independent experiments. **, P<0.01, ***, P<0.001, one-way ANOVA. shRNA against c-Met does not affect the number or total length of primary dendritic branches, but decreases the complexity of dendritic arborization shRNA against c-Met decreases synaptic clustering of NR2B and PSD-95 Figure 3. A. Cultured hippocampal neurons co-transfected with either shRNA vector or c-Met shRNA and GFP constructs were immunostained against GFP, NR2B and PSD-95. B. Quantification of NR2B and PSD-95 cluster sizes.C. Cumulative frequency plot showed that c-Met shRNA shifts the curve towards smaller of NR2B clusters. Error bars represent SEM. n>500 clusters per condition from three independent experiments. ***, P<0.001, one-way ANOVA. Treatment of neurons with PHA-665752 blocks HGF-induced synaptic clustering of NR2B Figure 7.A. Cultured hippocampal neurons were co-transfected with either shRNA Vector, scrambled shRNA (shRNA ScrM), or c-Met shRNA and GFP-Actin constructs. Cells were then fixed and immunostained against GFP. B, C. The number (B) or total length (C) of primary dendrites was quantitatively analyzed and shown in bar graphs. D1, D2, D3, D4. As shown in Sholl analysis, c-Met shRNA significantly reduced the complexity of dendritic branches (D1, D2, D3), while there was no significant difference among the three control conditions (D4). Error bars represent SEM. n>30 neurons per condition from three independent experiments. *, P<0.05, **, P<0.01, ***, P<0.001, two-way ANOVA. REFERENCES Figure 4. A. Synaptic NR2B clusters were enlarged by HGF treatment. However, pretreatment of neurons with PHA-665752 blocked the increase in size of NR2B in response to HGF. B. Quantification of NR2B cluster size. C. Cumulative analysis of the effect of HGF and PHA-665752 on synaptic NR2B clustering. Error bars represent SEM. n>500 clusters per condition from three independent experiments. ***, P<0.001, one-way ANOVA. 1. Campbell DB, Sutcliffe JS, Ebert PJ, et al. (2006) A genetic variant that disrupts MET transcription is associated with autism. Proc Natl Acad Sci. 103: 16834-16839. 2. Campbell DB, D'Oronzio R, Garbett K, Ebert PJ, Mirnics K, Levitt P, Persico AM. (2007) Disruption of cerebral cortex MET signaling in autism spectrum disorder. Ann Neurol. 62: 243-250. 3. Tyndall SJ & Walikonis RS. (2006) The receptor tyrosine kinase Met and its ligand hepatocyte growth factor are clustered at excitatory synapses and can enhance clustering of synaptic proteins. Cell Cycle. 5: 1560-1568. 4. Tyndall SJ, Patel SJ, Walikonis RS. (2007) Hepatocyte growth factor-induced enhancement of dendritic branching is blocked by inhibitors of N-methyl-D-aspartate receptors and calcium/ calmodulin-dependent kinases. Journal of Neuroscience Research. 85:2343–2351. 5. Lim CS & Walikonis RS. (2008) Hepatocyte growth factor and c-Met promote dendritic maturation during hippocampal neuron differentiation via the Akt pathway. Cellular Signalling. 20: 825-835. CONCLUSIONS Defects in signaling by c-Met could potentially impair neural connectivity in the following ways: • Reducing dendritic spine size and density. • Altering the balance between excitatory and inhibitory synapses. • Reducing dendritic complexity.