Download

1 / 23
250 likes | 555 Views
Single Molecule Biophysics. Reading : van Holde Chapter 16 Homework : due Wednesday, April 9 (in class) Van Holde: 16.1, 16.2, 16.3, 16.4 (consult original papers) Graduate students: Please schedule presentations Remember: Project papers are due last day of class (April 30). Overview:

E N D
Single Molecule Biophysics Reading: van Holde Chapter 16 Homework: due Wednesday, April 9 (in class) Van Holde: 16.1, 16.2, 16.3, 16.4 (consult original papers) Graduate students: Please schedule presentations Remember: Project papers are due last day of class (April 30) • Overview: • Why single-molecule biophysics? • Single-molecule fluorescence, TIRF illumination & applications • Atomic Force Microscopy (AFM) & applications (AFM imaging, AFM single-molecule force measurements) • Optical tweezers (laser tweezers, laser traps) & applications
Single molecule biophysics – Why? Quick example: The graph shows the speed of a population of polymerase molecules. What is the approximate, ensemble-average speed (as measured by bulk experiments)? What additional information can you obtain from single molecules? • Observation and physical manipulation of single, dynamic biomolecules. • A recently emerged, new field offering much deeper analysis of molecular behavior. • Most previous studies have been on the average, ensemble behavior of molecules. • Why study single molecules? • How any one molecule behaves is not revealed in bulk studies. • An individual enzyme may exist in two or more states of activity that are not revealed by bulk studies. (e.g speed of RNA polymerase transcription). • Analogy: How insurance actuary and novelist look at human behavior. • Static heterogeneity: Different enzyme molecules function at different rates (e.g. a lame population and a fast population) • Dynamic heterogeneity: A given single enzyme can switch between different rates. • Can take force measurements (force spectroscopy) on single molecules (previously impossible). E.g. motor proteins, unfolding-refolding proteins
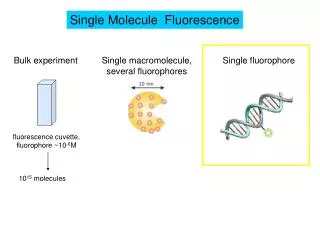
Single molecule fluorescence • Like Chapter 11 (bulk fluorescence), except on a singlefluorophores • Challenges: • Photobleaching: Most fluorophores photobleach, i.e. after absorbing many photons ( usually a few million; one photon is one absorption/emission cycle), they chemically rearrange and stop fluorescing (end of experiment). • Signal to noise ratio (need to reduce noise and have sensitive detection). • Must have very, very clean sample. • Detecting weak signals requires very sensitive instrumentation. • Needed: • A strong fluorophore (high absorbance and Q-yield). • Intense light source (often laser). • Very sensitive camera/detector • Eliminate all background light. • Illuminate a very small region only • Confocal microscopy (images only slices (stacks) of a sample) • Use TIR (total internal reflection).(On white board): From Snell’s law : When light encounters a lower index of refraction medium the light gets totally reflected when the incident angle is larger than the critical angle. We get an evanescent wave (evanescent = tending to vanish like vapor).
Prism-based TIR (total internal reflection) illumination Distance above cover slip Relative light intensity 132 nm 0.125 No signal (background) from molecules far from surface. 88 nm 0.25 Laser 44 nm 0.5 Only molecules close to surface will fluoresce Microscope objective Prism and cover slip TIRF No TIRF n1 …index of refraction of glass slide n2 …index of refraction of water q … incident angle l … wavelength of light • TIRF is a technique to only excite fluorophoses on/close to surface • Background noise is strongly reduced L. Peng et al. Microscopy Research & Technique (2007) 70, 372-81
Objective-based TIRF illumination Ray paths (schematic): 5 Total reflection1: Objective, 2: Immersion oil n = 1.518, 3: Cover slip n = 1.518, 4: Evanescent field, 5: Mountant n = 1.33…1.38 Angle of incidence smaller than the critical angle. www.zeiss.com See applet and info at: http://www.olympusmicro.com/primer/techniques/fluorescence/tirf/tirfhome.html
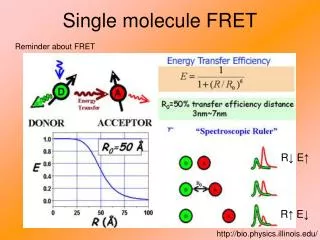
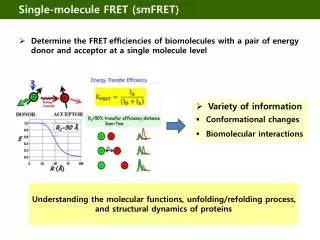
Application: Single molecule FRETG. Bokinsky et al. “Single-molecule transition-state analysis of RNA folding” (2003) PNAS 100: 9302 The hairpin ribozyme: The docking and undocking conformations and a single molecule FRET trace: SA
Application: Single molecule FRETG. Bokinsky et al. “Single-molecule transition-state analysis of RNA folding” (2003) PNAS 100: 9302 • From figure on previous page: • The ribozyme exhibits fluctuations between two states, docked and undocked FRET signal changes abruptly when a transition occurs. • Rate constants can be extracted from dwell times!! • They found a single rate constant for docking kdock = 0.018 sec-1. • They found four different rate constants for undocking • (ku = 0.01, 0.1, 0.8, 6 sec-1), which would have been hard to find in bulk studies). For single rate. Occurrence 1/e = 37% max t On-time, docked time (msec)
Schematic of an AFM Photodetector Laser Cantilever Piezo-electric transducer Sample Substrate Force controlled by feedback Images from NT-MDT web page
Atomic Force Microscopy • Advantages: • Can achieve atomic resolution on hard, crystalline surfaces. • Can often achieve nanometer resolution on biological samples. • Imaging can be done in buffer can image (biological) processes. • Can also be used to mechanically manipulate molecules (more in a bit). Gold surface (atomic resolution) S. Dutta, M. J. Snyder, D. Rosile, K. L. Binz, E. H. Roll, J. Suryadi, U. Bierbach, M. Guthold “PT-ACRAMTU, A Platinum–Acridine Anticancer Agent, Lengthens and Aggregates, but does not Stiffen or Soften DNA”, Cell Biochemistry and Biophysics (2013), DOI 10.1007/s12013-013-9614-8 (11 pages)
Tapping 4 nm On Off On Off 0 nm On Off On 2 mm scan Contact mode AFM scan Tapping mode AFM Contact mode (constant force mode): Use cantilever deflection as feedback signal, (use softer cantilevers, can still have lateral forces pushing molecules around) Tapping mode: oscillated cantilever at its resonance frequency (10 kHz to 300 kHz), use cantilever amplitude or phase as feedback signal. (Lateral forces mostly eliminated).
Transcribing RNA Polymerase Imaged by AFM1,2 100 nm T = 0 s T = 80 s T = 130 s T = 170 s A B C D T = 210 s T = 250 s T = 290 s 5.0 nm E F G 0 nm H Kasas movie 1. Kasas et al. (1997) Biochemistry 36(3), 461-468 2. Guthold et al. (1999) Biophys. J. 77, 2284-2294
Normal force measurements. Example 1. Protein unfolding Force: F = k·Dx; k … spring constant of cantilever Dx … deflection of cantilever (a) The principal AFM components. (b) Mechanical unfolding of repeating immunoglobulin-like domains (1). As the distance between the surface and tip increases (from state 1 to state 2), the molecule extends and generates a restoring force that bends the cantilever. When a domain unfolds (state 3), the free length of the protein increases, relaxing the force on the cantilever. Further extension again results in a restoring force (state 4). The last peak represents the final extension of the unfolded molecule before detachment from the SFM tip (state 5). • Carrion-Vazquez et al. “Mechanical and chemical unfolding of a single protein: a comparison” (1999) PNAS 96 3694-99 • Figure from Bustamante, Macosko, Wuite “Grabbing the cat by the tail: Manipulating molecules one by one. Nature reviews Molecular Cell Biology1 131-6
Normal force measurements. Example 2. Ligand binding forces and how they related to the koff rate (force spectroscopy). • Protein-ligand is spanned between the tip and the substrate. • The tip is then retracted, and, thus, applying a force to the bonds under investigation. • If the force is measured as a function of the pulling rate, it is termed force spectroscopy. Figure from: Guthold, M., Superfine, R., Taylor, R. (2001). The rules are changing: Force measurements on single molecules and how they relate to bulk kinetics. Biomedical Microdevices, 3, 9-18
Connection between rupture force and off-rate k-1 Bell model: an applied force lowers the activation energy. Assume a two-state model for the reaction. Dissociation rate without an applied force: Dissociation rate with applied force: G. Bell (1978) Science200, 616-627; E. Evans & K Ritchie (1997) Biophys. J.72, 1541-55
Connection between rupture force and off-rate, k-1 Experiment: Measure rupture force as a function of pulling rate. (here done with two different proteins). For this treatment, we assume the reaction proceeds far from equlibrium. The faster you pull the higher the rupture force. The rupture force is related to the off-rate F … rupture force T … temperature k-1 … off-rate x-1 … width of potential kB …Boltzmann constant Data from F. Schwesinger et al. (2000) PNAS97, 9972-77, First done by Rief at al. Science (1997) 276, 1109-12
How does a laser trap work? • Light “consists of photons, which carry momentum. Momentum is conserved. When light is absorbed, reflected or refracted, tiny forces on the order of piconewtons are generated. • For a laser trap we need a light gradient (light is focused). • Opposing scattering (down) and gradient forces (up), trap a bead in the focus. • If moved from the focus, bead is pulled back toward the focus. Laser beam lens Bead is below center of focus force on bead toward focus • Particle radius has to be larger than wavelength of light (Mie scattering regime). • Need transparent dielectric bead with index of refraction larger than surrounding medium. Fscatt. Fgrad
Ray-diagram for a bead to the left and higher than laser focus Restoring force of a laser trap A trap exerts a linear restoring force proportional to trap stiffness (force is linear to displacement). Using optical tweezers, one can apply pico-newton sized loads and measure nanometer level displacements. From Hubmayr lab, Mayo Clinic): http://mayoresearch.mayo.edu/mayo/research/hubmayr/
Laser tweezers • Force clamp: Force on molecule is kept constant by always having bead at the same position in laser trap ( feedback loop moving bead or stage). • Position clamp: Position of molecule is kept constant bead is pulled out of trap and, thus, force increases.
Applications of laser tweezers • Transcription by single RNA polymerase. Set-up for measuring force-velocity relation of a transcribing RNA polymerase (Wang et al. (1998) Science282, 902-7. Stall force is about 20 pN.
From Block lab: http://www.stanford.edu/group/blocklab/RNAP.html Applications of laser tweezers • Transcription by single RNA polymerase. Individual RNA polymerase molecules switch between a fast and a slow mode (Davenport et al. (2000) Science287, 2497-500 Dumbbell set-up used for some experiments
Mechanical properties of DNA • Stretching of double-stranded l-phage DNA; Length for B DNA ~ 16 mm). • Up to a length of about 15 mm: worm-like chain is straightened (entropy), little force needed. • Steep part of curve corresponds to elastic stretching of extended chain. • At ~ 17 mm a major conformational change occurs conversion to S-DNA (stretched-DNA). • Then DNA denatures and becomes single-stranded. From Smith et al. (1996) Science271, 795
A few more applications • Force-producing properties of various molecular motors, including kinesin moving along microtubules 13, actomyosin complexes 28, RNA polymerase 36, 48 and DNA polymerase 11 (Fig. 4b). Mechanical properties of DNA 47 • Mechanical unfolding of proteins 52, 53 • Unfolding of RNA molecules (54) • For a somewhat recent review see: Bustamante et al. “Grabbing the cat by the tail: Manipulating molecules one by one.” (2000) Nature reviews Molecular Cell Biology1 131-6
11. Wuite, G. J., Smith, S. B., Young, M., Keller, D. & Bustamante, C. Single-molecule studies of the effect of template tension on T7 DNA polymerase activity. Nature404, 103-106 (2000). | Article | PubMed | ISI | ChemPort | • 13. Svoboda, K., Schmidt, C. F., Schnapp, B. J. & Block, S. M. Direct observation of kinesin stepping by optical trapping interferometry. Nature365, 721-727 (1993). | Article | PubMed | ISI | ChemPort | • 28. Essevaz-Roulet, B., Bockelmann, U. & Heslot, F. Mechanical separation of the complementary strands of DNA. Proc. Natl Acad. Sci USA94, 11935-11940 (1997). | Article | PubMed | ChemPort | • 47. Smith, S. B., Cui, Y. & Bustamante, C. Overstretching B-DNA: the elastic response of individual double-stranded and single-stranded DNA molecules. Science271, 795-799 (1996). | PubMed | ISI | ChemPort | • 48. Yin, H. et al. Transcription against an applied force. Science270, 1653-1657 (1995). | PubMed | ISI | ChemPort | • 52. Kellermayer, M. S., Smith, S. B., Granzier, H. L. & Bustamante, C. Folding-unfolding transitions in single titin molecules characterized with laser tweezers. Science276, 1112-1116 (1997); erratum 277, 1117 (1997). | PubMed | ISI | ChemPort | • Tskhovrebova, L., Trinick, J., Sleep, J. A. & Simmons, R. M. Elasticity and unfolding of single molecules of the giant muscle protein titin. Nature387, 308-312 (1997). | Article | PubMed | ISI | ChemPort | • (J. Liphardt et al. “Reversible Unfolding of Single RNA Molecules by Mechanical Force” (2001) Science292, 733-737