Download
1 / 1
10 likes | 239 Views
*. *. *. *. *. *. *. *. *. *. 65. Female. 60. Male. 55. 50. Correct judgements (%). 45. Mean percentage of accuracy. 40. 35. 30. Male-visual priming. Female-visual priming. Motor priming. Neutral priming. Neutral priming. Male-visual priming.

E N D
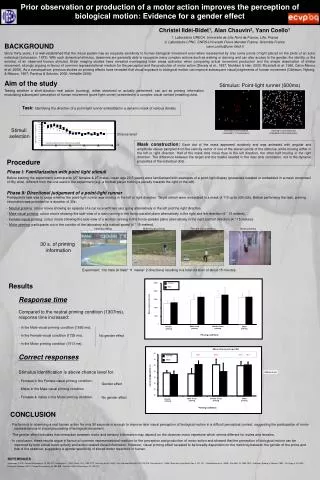
* * * * * * * * * * 65 Female 60 Male 55 50 Correct judgements (%) 45 Mean percentage of accuracy 40 35 30 Male-visual priming Female-visual priming Motor priming Neutral priming Neutral priming Male-visual priming Female-visual priming Priming conditions Priming conditions BACKGROUND Since thirty years, it is well-established that the visual system has an exquisite sensitivity to human biological movement even when represented by only some points of light placed on the joints of an actor individual (Johansson, 1973). With such dynamical stimulus, observers are generally able to recognize many complex actions such as walking or dancing and can also access to the gender, the identity, or the emotion of an observed human stimulus. Brain imaging studies have revealed overlapping brain areas activation when comparing actual movement production and the simple observation of similar movement, strongly arguing in favour of common representational medium for the perception and the production of motor action (Decety et al., 1997; Nishitani & Hari, 2000; Rizzolatti et al. 1996, Calvo-Merino et al. 2005). As a consequence, previous studies on priming effects have revealed that visual exposure to biological motion can improve subsequent visual judgements of human movement (Olofsson, Nyberg, & Nilsson, 1997; Pavlova & Sokolov, 2000; Verfaillie 2000) Aim of the study Testing whether a short-duration real action (running), either observed or actually performed, can act as priming information modulating subsequent perception of human movement (point-light runner) presented in a complex visual context (masking dots). Stimulus: Point-light runner (600ms) Task: Identifying the direction of a point-light runner embedded in a dynamic mask of various density Stimuli selection Point-light runner facing left embedded in 11 masking dots) Point-light runner facing left embedded in 220 masking dots) Chance level Mask construction: Each dot of the mask appeared randomly and was animated with angular and amplitude values sampled from the velocity vector of one of the eleven points of the stimulus, while moving either in the left or right direction. Half of the mask dots move thus in the left direction, the other half moving in the right direction. The difference between the target and the masks resided in the inter-dots correlation, not in the dynamic properties of the individual dots. *: statisticaly different from chance level, p<0.05 110 Step=11 dots 220 Prior observation or production of a motor action improves the perception of biological motion: Evidence for a gender effect • Procedure • Phase I: Familiarization with point light stimuli • Before starting the experiment, participants (27 females & 27 males, mean age 20.5 years) were familiarised with examples of a point-light display (presented isolated or embedded in a mask composed of fifty dots), different from the one used in the experiment (e.g. a football player kicking a penalty towards the right or the left). • Phase II: Directional judgement of a point-light runner • Participant’s task was to judge whether the point-light runner was moving in the left or right direction. Target stimuli were embedded in a mask of 110 up to 220 dots. Before performing the task, priming information was provided for a duration of 30s: • Neutral priming: colour movie showing an episode of a car race with two cars going alternatively in the left and the right direction. • - Male-visual priming: colour movie showing the side-view of a man running in the fronto-parallel plane alternatively in the right and left direction (4 * 15 meters). • - Female-visual priming: colour movie showing the side-view of a woman running in the fronto-parallel plane alternatively in the right and left direction (4 * 15 meters). • - Motor priming: participants run in the corridor of the laboratory at a natural speed (4 * 15 meters). Neutral priming Male-visual priming Female-visual priming Motor priming 30 s. of priming information Experiment: 132 trials (6 trials* 11 masks* 2 directions) resulting in a total duration of about 15 minutes. Christel Ildéi-Bidet1, Alan Chauvin2, Yann Coello1 1. Laboratoire URECA, Université de Lille Nord de France, Lille, France 2. Laboratoire LPNC, CNRS-Université Pierre Mendes France, Grenoble France yann.coello@univ-lille3.fr Results 2500 Female Male 2000 • Response time • Compared to the neutral priming condition (1307ms), response time increased: • In the Male-visual priming condition (1592 ms). • - In the Female-visual condition (1725 ms). • In the Motor priming condition (1513 ms). 1500 Mean response time (ms) 1000 500 0 Male-visual priming Female-visual priming Motor priming Neutral priming No gender effect Priming conditions Above chance level (p<0.05) 56% 56% 54% 55% • Correct responses • Stimulus identification is above chance level for: • Females in the Female-visual priming condition. • - Males in the Male-visual priming condition. • - Females & males in the Motor priming condition. Chance level Gender effect No gender effect • CONCLUSION • Performing or observing a real human action for only 30 seconds is enough to improve later visual perception of biological motion in a difficult perceptual context, suggesting the participation of motor • representations in visual processing of biological movement. • The gender effect indicates that interaction between motor and sensory information may depend on the observer motor repertoire which seems different for males and females. • In conclusion, these results argue in favour of common representational medium for the perception and production of motor action and showed that the perception of biological motion can be • improved by both actual motor activity and action-related visual information. However, visual priming effect revealed to be broadly dependent on the matching between the gender of the prime and • that of the observer, suggesting a gender specificity of stored motor repertoire in human. REFERENCES Johansson (1973). Percept Psychophys, 14, 201-211 ; Decety et al., (1997). Brain, 120, 1763-1777 ; Nishitani & Hari (2000). Proc Nat Acad Sci-USA, 97, 913-918 ; Rizzolatti et al. (1996). Brain Res Cogn Brain Res, 3, 131-141 ; Calvo-Merino et al. (2005). Curr Biol, 16, 1905-1910 ; Olofsson, Nyberg, & Nilsson (1997). Vis Cogn, 4, 373-382 ; Pavlova & Sokolov (2000). Percept Psychophys, 62, 889-899 ; Verfaillie (2000). Brain Cogn, 44, 192-213.