第三章: DNA 复制 ( DNA Replication)
第三章: DNA 复制 ( DNA Replication). 第一节: DNA 复制的基本原则. 一、 Semiconservative Replication of Double-Strand DNA. 概 念.

第三章: DNA 复制 ( DNA Replication)
E N D
Presentation Transcript
概 念 在复制过程中首先两条亲本链之间的氢键断裂,双链分开,然后以每条亲本链(parental strand)为模板,按碱基互补配对原则(A:T,G:C),由DNA聚合酶催化合成新的互补子链(daughter strand)。复制结束后,每个子代DNA的一条链来自亲代DNA,另一条链则是新合成的,并且,新形成的两个DNA分子与原来的DNA分子的碱基序列完全相同。这种复制方式称为半保留复制(semiconservation replication)。
二、DNA复制的化学 DNA合成的底物是脱氧核苷5’-三磷酸(dNTP):dATP,dGTP,dCTP和dTTP。复制的机制涉及到生长链的3’-OH对与模板上的碱基互补的dNTP的α-磷酸基团进行亲核进攻,释放出焦磷酸。聚合反应所需的能量来源于dNTP和焦磷酸的水解。
三、复制起点与复制子 Replicon:作为一个单位进行复制的任何一段DNA序列。 它含有一个复制起点,有时还含有一个复制终点。 Origin:是复制子起始复制的一段DNA序列。
作为一个单位进行复制的任何一段DNA序列。 复制子的复制通常从一个固定的位点开始的,这种起始DNA复制的序列叫做复制起点。DNA复制时,双螺旋的两条链在复制起点处分开形成两条模板链。一旦复制开始,就会在DNA分子上形成两个复制叉(replication fork)。复制叉向着未复制的它们沿着DNA相向移动,因此复制是双向的。所有的原核生物的染色体,很多噬菌体和病毒的DNA分子是环状的,它们作为单个复制子完成复制。
真核生物染色体的线状DNA则含有多个复制子,每一个复制子都有自己的复制起点。一个典型的哺乳动物细胞有50 000-100 000个复制子,复制子的长度为40-200 Kb。正在复制的真核生物基因组DNA分子上,会形成许多复制泡。随着复制叉沿着DNA分子向两个方向移动,复制泡不断变大,最终,两个相邻复制泡的复制叉会相遇、融合,完成DNA的复制。
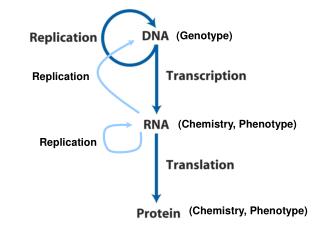
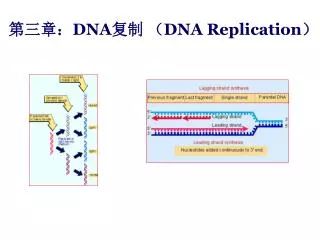
在复制叉处,DNA的两条链都作为模板同时合成两条新链。DNA分子的两条链是反向平行的,而DNA复制时无论以那条链做模板,新合成的链是按5’→3’方向进行的,所以只有一条模板链指导合成的新生链能够沿着复制叉运动的方向连续复制,此新合成的链称为前导链(leading strand)。另一条模板链指导合成的新生链也是沿5’→3’方向进行,但与复制叉前进的方向相反,是分段合成的,这些片断于1969年首先在大肠杆菌中分离出来,被称为冈崎片段。
在细菌中冈崎片段长约1000-2000 核苷酸,但在真核生物中,相应片断的长度可能短得多,由100~400个核苷酸组成。合成的片段即为冈崎片段。这些冈崎片段以后由DNA连接酶连成完整的DNA链,因此冈崎片段是DNA复制中短暂的中间产物。这条链不连续合成的链称为滞后链(lagging strand) 。这种前导链的连续复制和滞后链的不连续复制在生物界是普遍存在的,称为DNA合成的半不连续复制。
Other replicated in short segments(to be joined later by ligase) One strand replicated as continuous segment P C o n t i n u o u s S Unzipping P S e g m e n t e d ReplicationFork S P S P S
五、RNA引导 目前已知的DNA聚合酶都只能延伸已经存在的DNA链,而不能从头启动DNA链的合成,这是因为它在合成DNA时需要一个自由的3’ OH。那么一个新的DNA链的合成是如何开始的呢?研究发现,DNA复制时还需要另外一种酶来合成一段RNA作为合成DNA的引物。在细菌中,引物合成依靠引物酶(primase),它是一种与转录酶不相关的RNA聚合酶。
一、复制的起始 1. 大肠杆菌的复制起点(OriC) 大肠杆菌的复制起点称为OriC。 将大肠杆菌的染色体随机断裂后插入到缺乏复制起点的质粒中,由于质粒上含有选择标记,凡能存活的克隆,质粒中都含有一段控制DNA复制的序列。通过缺失分析鉴定出大肠杆菌的复制起点约有240 bp组成。
该序列含有两个短的重复基序,两个基序的长度分别为9 bp和13 bp。9 bp基序共有4个拷贝,分散于整个OriC的范围内,它是DnaA蛋白的结合位点。通过DnaA蛋白与OriC的相互作用以及DnaA蛋白之间的相互作用,大约30个DnaA蛋白结合在复制起始区。这种结合只能发生在负超螺旋DNA分子上,而负超螺旋是大肠杆菌染色体的正常存在状态。
DNA分子在DnaA蛋白复合体上的缠绕,导致双螺旋在串连排列的3个富含AT的13 bp基序处解开。一般认为螺旋解开是由于DNA双螺旋在DnaA蛋白复合体上缠绕后产生的扭转张力的结果。
Helicase (DnaB) continues to separate strands DNA双螺旋的进一步打开需要DnaB蛋白。DnaB蛋白是解旋酶(Helicase),其作用是解开DNA双链。两个DnaB蛋白在复制起点处分别与两条单链结合,并按5’→3’相向而行打开DNA双链。在复制起点处DNA双链打开以后,DnaA蛋白便会募集由6个DnaB和6个DnaC构成的复合体与起始位点处的单链DNA结合。DnaC为DnaB蛋白装载因子,依靠DnaC蛋白的单链DNA结合域以及与DnaA蛋白的相互作用,DnaB和DnaC蛋白复合体结合在复制起始位点。
然后,DnaC催化DnaB蛋白解环,并套在单链DNA上。在DnaB和DnaC蛋白复合体中,DNA解旋酶保持非活性状态。DNA解旋酶的安装导致装载因子从复合体中释放出来,并激活DNA解旋酶。DNA解旋酶向前运动,在其身后留下单链DNA模板。然后,DnaC催化DnaB蛋白解环,并套在单链DNA上。在DnaB和DnaC蛋白复合体中,DNA解旋酶保持非活性状态。DNA解旋酶的安装导致装载因子从复合体中释放出来,并激活DNA解旋酶。DNA解旋酶向前运动,在其身后留下单链DNA模板。
接着单链结合蛋白(single-stranded binding protein, SSB)与解旋酶解开的单链结合,其作用是防止单链降解,阻止单链退火。SSB蛋白与单链DNA的结合有协同效应(cooperative),即一个SSB的结合会促进另一个SSB与单链DNA的结合。这种协同结合使单链DNA被解旋酶释放出后,很快被SSB所覆盖。一旦被SSB覆盖,单链DNA即处于伸直状态,有利于其作为模板进行DNA合成或RNA引物的合成。
二、Elongation 一旦复制起始,复制叉就沿着DNA分子前进,合成与亲本链互补的两条新DNA链。新生链的合成由DNA聚合酶催化完成。从大肠杆菌细胞中分离出了5种DNA聚合酶。DNA聚合酶III是大肠杆菌DNA复制中链延伸反应的主要聚合酶。DNA聚合酶I专门用于RNA引物的去处。另外3种DNA聚合酶主要参与DNA修复和跨损伤合成。
1.DNA聚合酶I DNA pol I由一条多肽链组成,分子量为109 KD。酶分子中含有一个Zn2+,是聚合酶活性必需的。用枯草杆菌蛋白酶可将此酶水解成两个片段,大片段的分子量为76 KD,通常称为klenow片段,小片段为34KD。大小片段具有不同的酶活性。
E. coli DNA pol I (polA) 1. easily cleaved into two fragments by proteinase: a. large fragment (Klenow fragment) contains polymerase and 3’-5’ exonuclease (proofreading domain). Used in vitro for synthesis reactions (DNAsequencing). Klenow fragment (68 kD) small fragment (35 kD) N C C exonuclease 3’-5’ polymerase site 5’-3’ activity proofreading catalytic exonuclease
这是DNA聚合酶最主要的活性,按模板DNA上的核苷酸顺序,将互补的dNTP逐个加到引物RNA3’-OH末端,即促进3'-OH与dNTP的5'-PO4形成磷酸二酯键,酶的专一性表现为新进入的dNTP必须与模板DNA碱基配对时才有催化作用,5'→3'聚合活性存在于klenow片段上。 DNA聚合酶I的延伸能力不强,每次与模板-引物结合仅能添加20~100个核苷酸。
(2)DNA聚合酶的3’→5’外切核酸酶 (exonuclease)活性 这种酶活性的主要功能是从3’ →5’方向识别并切除DNA生长链末端与模板DNA不配对的核苷酸,这种功能称为校对功能(proofreading),这是保证其聚合作用的正确性不可缺少的,因此对于维持DNA复制的真实性(replicational fidelity)至关重要。
(3)DNA聚合酶的5'→3'外切核酸酶活性 这种酶活性是从DNA链的5’端向3’末端水解已配对的核苷酸,本质是切断磷酸二酯键,每次能切除10个核苷酸。因此,这种酶活性在DNA损伤的修复中可能起重要作用,对完成的DNA片段去除5’端的RNA引物也是必须的。 DNA polⅠ的5'→3'聚合活性和5'→3'外切酶活性协同作用,可以使DNA一条链上的切口从5'→3'方向移动,这种反应叫做缺刻平移(nick translation)
2. DNA Polymerase III (DNA聚合酶 III) DNA聚合酶III由10个不同的亚基构成。其中α、ε和θ亚基构成核心酶。α亚基具有5'→3' 聚合酶活性,ε亚基具有3'→5'外切活性,θ亚基功能尚不清楚,可能仅仅起结构上的作用,使两个核心亚基以及其他各辅助亚基装备在一起。DNA聚合酶III具有两个拷贝的核心酶。
(Note: no beta subunits are shown; without beta, this form of the complex is called DNA pol III*)
两个拷贝的β亚基围绕DNA双螺旋形成一个环状的二聚体。一旦β亚基二聚体与DNA紧密结合,它的作用就像一个“滑动夹子”(sliding clamp)携带着核心聚合酶沿着DNA链自由滑动。这样,聚合酶的活性位点就可以一直定位在生长中的复制叉附近。另一方面,核心酶与滑动夹结合后,其延伸能力也显著提高。DNA聚合酶的延伸能力(processivity)定义为每次聚合酶与模板-引物结合时所能添加的核苷酸的平均数。与滑动夹的结合是如何改变DNA聚合酶的延伸能力的呢?在无滑动夹的情况下,DNA聚合酶平均每合成20~100个核苷酸就从DNA模板上脱落下来。
Beta forms a donut shaped ring around the DNA and helps to anchor the holoenzyme to the DNA during replication . By acting as a sliding "clamp", beta helps the holoenzyme to replicate long stretches of DNA without "falling off" the strand (this is called processivity).
但是,滑动夹可以阻止聚合酶脱落,从而大大增加了DNA聚合酶的延伸能力。聚合酶的第3个组成部分是由5个亚基构成的一个所谓的γ复合体。γ复合体又称夹子装载因子(clamp loader),催化滑动夹打开,并将其结合在引物-模板上。介导β亚基与模板-引物双螺旋的结合。最后,两个拷贝的τ亚基使核心聚合酶形成二聚体。
(Note: no beta subunits are shown; without beta, this form of the complex is called DNA pol III*)