Microscopy of Ribosome Structure and Function
30 likes | 280 Views
Microscopy of Ribosome Structure and Function. Justin Levy, Roger Ndindjock, James Potter, Justin Quon, Sam Veihmeyer . Current Mechanism of Translation. Early Microscopy

Microscopy of Ribosome Structure and Function
E N D
Presentation Transcript
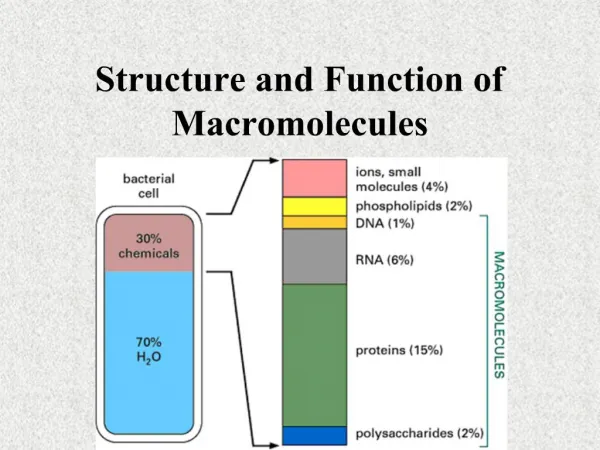
Microscopy of Ribosome Structure and Function Justin Levy, Roger Ndindjock, James Potter, Justin Quon, Sam Veihmeyer Current Mechanism of Translation Early Microscopy In the late 50’s, scientists began seriously considering the structure of the ribosome. Light microscopes had reached their theoretical limit twenty years prior at magnifications of 500X or 1000X. 10,000X magnification was needed to view organelle structures and much higher magnification was needed to visualize protein structure. The need for more advanced microscopy techniques was apparent. Neutron Diffraction Neutron Diffraction (ND) was developed in 1946 by Clifford G. Shull. In ND, neutrons are accelerated in a magnetic field and fired at the sample. Diffraction occurs when neutron waves (approximately 0.1 nm) encounter obstacles that are similar in size. The beam of neutrons is not affected by atomic electron clouds and will be diffracted only by atomic nuclei. In this regard, ND is superior to EM or X-Ray crystallography whose probes are affected by atomic electron density. Small-Angle-Neutron-Scattering (SANS) is a type of ND used for determining the structures of protein that relies on neutron scattering for detection. This technique was vital to the determination of tRNA locations in the mechanism for translation. SANS was also used to determine the ribosome’s atomic structure. 1 1 – Initiation factor binding 2 – Binding of tRNA with associated factors to form preinitiation complex 2 Electron Microscopy Since its initial development by Max Knoll and Ernst Ruska in 1931, electron microscopy (EM) has been of pivotal importance to the study of the ribosome. In EM, a beam of highly energetic electrons is directed at the sample to provide up to 500,000X magnification. Invaluable topological, morphological, compositional, and crystallographic insight can be obtained. 3 3 – eIF4G/eIF4A binding to preinitiation complex 4 4 – Association of preinitiation factor with mRNA Eukaryotic 80S Ribosome Initiation 40S Subunit 60S Subunit 5 5 – Small ribosome subunit scans for start codon 28S rRNA 5.8S rRNA 18S rRNA 5S rRNA 49 Ribosomal Proteins 33 Ribosomal Proteins 6 – Associated proteins falls off small subunit, large subunit attaches and translation begins 6 Large Subunit Atomic Structure (From http://en.wikipedia.org/wiki/Image:Ribosome_50s.png) (From http://www.emunix.emich.edu/~rwinning/genetics/pics/transl1.jpg) Common Types of Electron Microscopes: 1. Tunneling EM: Electrons pass through sample 2. Scanning EM: After intial bombardment, secondary electrons leave the sample to be detected. 3. Reflection EM: After incidence, reflected electrons are detected X-Ray Crystallography Australian William Henry Bragg and his son William Lawrence Bragghas won the 1915 Noble Prize in Physics for the invention of the X-Ray Crystallography (XRC) method. Due to difficulties in crystallization, ribosomes were not analyzed using XRC until the illuminating work by Yonath and Wittman in 1980. The 50S bacterial ribosome crystals prepared by Yonath and Wittman diffracted X-rays to 3.5 Å and provided scientists with a great deal of structural data. The structure of the 30S subunit was later revealed. In XRC, crystallized sample deflects X-rays to produce a diffraction pattern. This pattern corresponds to the number of atoms in the molecule. Heavier atoms scatter X-rays more effectively. Diffraction patterns are visualized from several angles to provide organizational information about the molecular components. Elongation 1 1 – Charged tRNAs (aminoacyl-tRNAs) bind to elongation factor eEF-1 in the presence of GTP 2 2 – The complex enters the empty A-site on a ribosome carrying an initiator Met-tRNA Cryo-Electron Microscopy Cryo-Electron Microscopy (CEM) is an EM technique that involves freezing biological samples in order to preserve the hydrated state of the specimen. This also provides protection in the high-radiation, low pressure environment during observation. Through CEM, binding sites of tRNA and elongation factors were elucidated. CEM also helped distinguish between the subunits of the ribosome. A key consideration in CEM is the formation of vitreous ice. Liquid ethane is used (-185°C) instead of liquid nitrogen (-195°C) because of increased heat capacity; N2(l) will boil off in the process of freezing, permitting unwanted ice crystals to form. 3 3 – After the correct codon-tRNA binding, the growing polypeptide in the P site is transferred to the A site. Peptidyl transferase attaches the amino acid to the nascent peptide chain 4 References Ban, Nissen, et al. “The Complete Atomic Structure of the Large Ribosomal Subunit at 2.4 Angstrom Resolution.” Science. 289. 2000. Frank, Joachim. “Toward an Understanding of the Structural Basis of Translation.” Genome Biology. 4:237. 2003. Moore, Peter. “The Ribosome at Atomic Resolution.” Biochemistry. 40 (11). 2001. Nierhaus, Wadzak, et al. “Structure of the elongating ribosome: Arrangement of the two tRNAs before and after translocation.” Proc. Natl. Acad. Sci. 95, 1998. 4 – Translocation occurs. The used tRNA leaves from the E site and the process repeats until a stop codon is reached Termination When the ribosome reaches a stop codon, release factors resembling tRNA bind to the A site and the nascent peptide chain is released