Download
1 / 1
10 likes | 119 Views
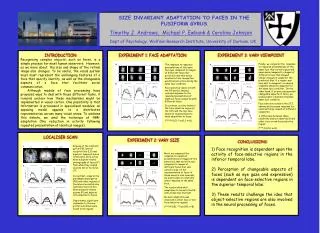
A. axial. coronal. sagittal. A. A. STS. STS. SIZE INVARIANT ADAPTATION TO FACES IN THE FUSIFORM GYRUS Timothy J. Andrews, Michael P. Ewbank & Caroline Johnson Dept of Psychology, Wolfson Research Institute, University of Durham, UK. PG. FG. LO. LO. FG. PG. MO. B. B. B. ***.

E N D
A axial coronal sagittal A A STS STS SIZE INVARIANT ADAPTATION TO FACES IN THE FUSIFORM GYRUS Timothy J. Andrews, Michael P. Ewbank & Caroline Johnson Dept of Psychology, Wolfson Research Institute, University of Durham, UK PG FG LO LO FG PG MO B B B *** * * C C C A B *** ** C INTRODUCTION Recognising complex objects, such as faces, is a simple process for most human observers. However, as we move about, the size and shape of the retinal image also changes. To be useful, the visual system must must represent the unchanging features of a face that specify identity, as well as the changeable aspects of a face that facilitate social communication. Although models of face processing have proposed ways to deal with these different tasks, it remains unclear how these mechanisms might be implemented in visual cortex. One possibility is that information is processed in specialised modules; an opposing model appeals to a distributed representation across many visual areas. To address this debate, we used the technique of fMR-adaptation (the reduction in activity following repeated presentation of identical images) . EXPERIMENT 1: FACE ADAPTATION EXPERIMENT 3: VARY VIEWPOINT Finally, we compared the response to repeated presentations of the same face that varied in gaze and expression compared to images of different faces that showed similar changes in viewpoint. We predicted that, if a region was invariant to viewpoint, a reduced response should be apparent for the same face condition . On the other hand, if an area represented changeable aspects of faces, we would expect a maximal response same face condition. Face selective voxels in the STS showed an increased response for identical faces viewed at different viewpoints. A difference between these conditions was not observed in any other face or non-face selective region. (***=P<0.01; n=8) The response to repeated presentations of the same face was compared to images of different faces Our prediction was that areas involved in face recognition would be less active during the same face condition Face selective voxels in both the FG and LO showed reduced response to the repeated presentation of identical faces compared with different faces. In contrast, activity levels in the face-selective region of the STS and the non-face selective regions failed to show adaptation to faces. (***=P<0.01; *=p<0.1; n=8) LOCALISER SCAN EXPERIMENT 2: VARY SIZE CONCLUSIONS 1) Face recognition is dependent upon the activity of face-selective regions in the inferior temporal lobe. 2) Perception of changeable aspects of faces (such as eye gaze and expression) is dependent on face-selective regions in the superior temporal lobe. 3) These results challenge the idea that object-selective regions are also involved in the neural processing of faces. Regions of the fusiform gyrus (FG), lateral occipital lobe (LO) and superior temporal sulcus (STS) were more active when subjects viewed photographs of faces (F) than when they viewed objects (O) or textures (T). In contrast, areas in the parahippocampal gyrus (PG) and other regions of the lateral occipital lobe (LO) were more active when subjects viewed scenes (P) and objects (O) compared to faces. Importantly, significant responses to the non-preferred stimuli were found in all regions. Next, we compared the response to repeated presentations of images of the same face that varied in size compared to images of different faces that also varied in size. If the representation of faces in these areas is size-invariant, we would expect a relatively lower response to the same face. The results show that adaptation to faces in the FG and LO was size invariant. No such adaptation was observed in other face or non-face selective regions. (***=P<0.01; **=p<0.05; n=8)





