
ΧΗΜΟΣΤΑΤΗΣ
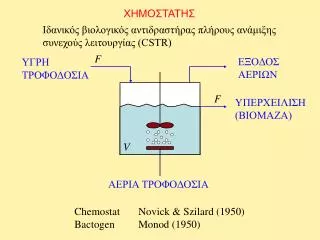
ΧΗΜΟΣΤΑΤΗΣ. Ιδανικός βιολογικός αντιδραστήρας πλήρους ανάμιξης συνεχούς λειτουργίας (CSTR). F. ΕΞΟΔΟΣ ΑΕΡΙΩΝ. ΥΓΡΗ ΤΡΟΦΟΔΟΣΙΑ. F. ΥΠΕΡΧΕΙΛΙΣΗ (ΒΙΟΜΑΖΑ). V. ΑΕΡΙΑ ΤΡΟΦΟΔΟΣΙΑ. Chemostat Novick & Szilard (1950) Bactogen Monod (1950). Ισοζύγιο βιομάζας. : ειδικός ρυθμός ανάπτυξης.

ΧΗΜΟΣΤΑΤΗΣ
E N D
Presentation Transcript
ΧΗΜΟΣΤΑΤΗΣ Ιδανικός βιολογικός αντιδραστήρας πλήρους ανάμιξης συνεχούς λειτουργίας (CSTR) F ΕΞΟΔΟΣ ΑΕΡΙΩΝ ΥΓΡΗΤΡΟΦΟΔΟΣΙΑ F ΥΠΕΡΧΕΙΛΙΣΗ (ΒΙΟΜΑΖΑ) V ΑΕΡΙΑ ΤΡΟΦΟΔΟΣΙΑ Chemostat Novick & Szilard (1950) Bactogen Monod (1950)
Ισοζύγιο βιομάζας : ειδικός ρυθμός ανάπτυξης D = F/V ρυθμός αραίωσης Αν xF = 0 (στείρα τροφοδοσία) σε μόνιμη κατάσταση: D =
παραγόμενη βιομάζα Ειδικός ρυθμός ανάπτυξης = (βιομάζα)(χρόνος) παραγόμενος αριθμός κυττάρων Ειδικός ρυθμός αναπαραγωγής n = (αριθμός κυττάρων)(χρόνος) logx n logn x t
Χημοστάτης με ένα περιοριστικό του ρυθμού θρεπτικό συστατικό (υπόστρωμα) στην τροφοδοσία sF x, s x, s Ισοζύγιο βιομάζας : Ισοζύγιο υποστρώματος: παραγόμενη βιομάζα Y : συντελεστής απόδοσης = καταναλισκόμενο υπόστρωμα
Ερώτημα: Το εξαρτάται και από το x ή μόνο από το s; Αν = f(s), τότεs = F(D) Αν = f(s,x), τότεs = F(D,sF) Πειράματα των Grady et al. (1972) - Aerobacter Aerogenes
Μοντέλο Monod (1942) Ισοζύγια σε χημοστάτη Σε μόνιμη κατάσταση
Τιμές συγκεντρώσεων σε μόνιμη κατάσταση Για να έχει φυσικό νόημα η μόνιμη κατάσταση πρέπει: x > 0, 0 < s < sF Παραγωγικότητα βιομάζας Μεγιστοποίηση παραγωγικότητας
x s P D
Παραγωγή προϊόντος Ισοζύγιο για το προϊόν: rp: ρυθμός παραγωγής προϊόντος rp= x προϊόν σχετιζόμενο με την ανάπτυξη rp= x προϊόν μη σχετιζόμενο με την ανάπτυξη [Luedeking & Piret (1959)] Παραγωγικότητα προϊόντος:Dp (μεγιστοποίηση;)
ΣΥΝΤΗΡΗΣΗ ΚΑΙ ΕΝΔΟΓΕΝΗΣ ΜΕΤΑΒΟΛΙΣΜΟΣ • Διατήρηση της διαφοράς των χημικών δυναμικών μεταξύ του εσωτερικού και του εξωτερικού των κυττάρων. • Ανασύνθεση μακρομορίων (πρωτεΐνες, νουκλεϊκά οξέα), που έχουν αποσυντεθεί στα μονομερή τους. • Κίνηση των κυττάρων.
Ενδογενής μεταβολισμός Ισοζύγιο βιομάζας: [Herbert (1958)] Σε μόνιμη κατάσταση: x D
Φαινόμενος συντελεστής απόδοσης κλίση D
Συντήρηση Ισοζύγιο υποστρώματος: [Marr et al. (1963), Pirt (1966)] Φαινόμενος συντελεστής απόδοσης κλίση D
ΑΝΤΙΔΡΑΣΤΗΡΑΣ ΗΜΙΔΙΑΛΕΙΠΟΝΤΟΣ ΕΡΓΟΥ F V • Προσθήκη ρυθμιστικών ουσιών • Διατήρηση χαμηλών συγκεντρώσεων υποστρωμάτων για αποφυγή παρεμποδιστικών φαινομένων • Επιμήκυνση της στατικής φάσης ανάπτυξης μικροοργανισμών
Συνολικό ισοζύγιο μάζας αν r = const. Ισοζύγιο συστατικού
Ανάπτυξη με παρεμπόδιση από το υπόστρωμα s
V(t) F(t) t x(t) s(t) t
Μικροβιακή ανάπτυξη σε ιδανικό αυλωτό αντιδραστήρα
ΜΗ ΙΔΑΝΙΚΟΙ ΒΙΟΑΝΤΙΔΡΑΣΤΗΡΕΣ • Ατελής ανάμιξη • Απώλεια υγρού λόγω εξάτμισης • Ασυνεχής τροφοδοσία • Αφρισμός, συσσωμάτωση και παράσυρση μικροοργανισμών στην επιφάνεια • Προσκόλληση μικροοργανισμών και ανάπτυξη στα τοιχώματα
Ατελής ανάμιιξη Κατανομή χρόνων παραμονής ΑΝΤΙΔΡΑΣΤΗΡΑΣ
κλάσμα υγρού στο ρεύμα τροφοδοσίας που παρέμεινε στον αντιδραστήρα για χρόνο t ως t+dt η k ροπή της E(t) Μέσος χρόνος παραμονής
Κατανομή χρόνων παραμονής ιδανικού χημοστάτη 1 D Μέσος χρόνος παραμονής
Κατανομή χρόνων παραμονής ιδανικού αυλωτού αντιδραστήρα 1 Μέσος χρόνος παραμονής
Διασπορά χρόνων παραμονής όπου Ιδανικός CSTR: Ιδανικός αυλωτός αντιδραστήρας:
Μοντέλα αντιδραστήρων ατελούς ανάμιξης Ισοζύγια σε μόνιμη κατάσταση Τμήμα 1 Τμήμα 2
Αν Bailey & Ollis (1986)
Ν χημοστάτες Ισοζύγια σε μόνιμη κατάσταση Βιομάζα Υπόστρωμα i = 1,…,N Αν και Με ανακύκλωση:
Αυλωτός αντιδραστήρας με αξονική διάχυση Αν ή Συνοριακές συνθήκες:
Προσκόλληση κυττάρων στα τοιχώματα του αντιδραστήρα • Σχηματισμός στρώματος (film) από προσκόλληση των κυττάρων μεταξύ τους. • Σχηματισμός μονοκυτταρικής στοιβάδας. Τα κύτταρα προσκολλώνται στο τοίχωμα, αλλά όχι και μεταξύ τους.
Μοντέλο Topiwala & Hamer (1971)Υποθέσεις • H πυκνότητα των κυττάρων που είναι προσκολλημένα στα τοιχώματα είναι σταθερή. • H ανάπτυξη των κυττάρων στα τοιχώματα αντισταθμίζεται από την είσοδο κυττάρων στο υγρό. • Tα κύτταρα που είναι προσκολλημένα στα τοιχώματα δεν υφίστανται αποκόλληση. • O ρυθμός ανάπτυξης επάνω στα τοιχώματα είναι ίδιος με εκείνον στο υγρό.
Εξισώσεις μοντέλου Σε μόνιμη κατάσταση:
x P D
Γενίκευση του μοντέλου (Baltzis & Fredrickson, 1983)
x D