Download
1 / 45
450 likes | 605 Views
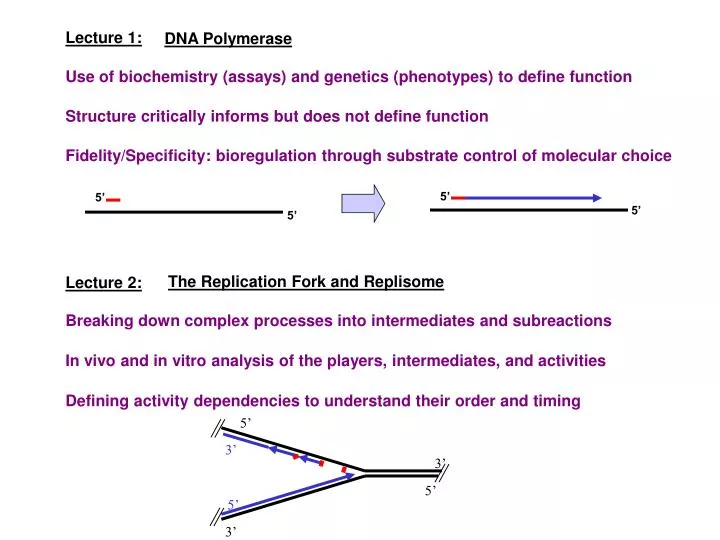
5’. 3’. 3’. 5’. 5’. 3’. Lecture 1:. DNA Polymerase. Use of biochemistry (assays) and genetics (phenotypes) to define function. Structure critically informs but does not define function. Fidelity/Specificity: bioregulation through substrate control of molecular choice. 5’. 5’. 5’. 5’.

E N D
5’ 3’ 3’ 5’ 5’ 3’ Lecture 1: DNA Polymerase Use of biochemistry (assays) and genetics (phenotypes) to define function Structure critically informs but does not define function Fidelity/Specificity: bioregulation through substrate control of molecular choice 5’ 5’ 5’ 5’ The Replication Fork and Replisome Lecture 2: Breaking down complex processes into intermediates and subreactions In vivo and in vitro analysis of the players, intermediates, and activities Defining activity dependencies to understand their order and timing
Synthesis with discontinuous lagging strand pieces (okazaki fragments) requires repeated: A. priming B. replacement of primed sequence C. ligation Okazaki fragment length: prokaryotes 1000 - 2000 nt; eukaryotes 100-200 nt Semi-Discontinuous DNA Synthesis at Replication Forks Leading daughter strand: polymerase moves continuously in same direction as replication fork Lagging daughter strand: polymerase moves discontinuously in opposite direction as replication fork 5’ Fork Movement C 3’ B lagging A 3’ 5’ leading 5’ 3’
Understanding Molecular Mechanisms S P S = substrate From This: P = product A1 A2 A3 A4 An+1 I = intermediates I4………….. In S I1 I2 I3 P To This: A = activity The transformation of one intermediate into another defines activities which are performed by proteins and/or nucleic acids How does one detect and identify intermediates? How does one structurally characterize intermediates? How does one identify the proteins/nucleic acids responsible for the activities?
Understanding Molecular Mechanisms S P S = substrate From This: P = product A1 A2 A3 A4 An+1 I = intermediates I4………….. In S I1 I2 I3 P To This: A = activity The transformation of one intermediate into another defines activities which are performed by proteins and/or nucleic acids How does one identify and detect intermediates? How does one structurally characterize intermediates? How does one identify the proteins/nucleic acids responsible for the activities?
I1 I1 I2 I2 P P Detecting Intermediates The Challenge: Intermediates are often transient, scarce, and coexist with other molecular species P S I1 I2 Synchronize Reaction To Transiently Enrich Intermediates Pulse-Chase Label Synchronous Cohort Block Reaction Step To Accumulate Intermediate S P S P S I1 I2 I1 I2 I1 I1 P P S { I2 I2 S I2 P S I1 I1 P Partial Reaction P S I1 I2 I2 Examples of blocks: Can provide temporal order of intermediates Can provide temporal order of intermediates Remove or inactivate protein Remove cofactor Label can sensitivity of detection Single molecule analyses follow this principal but don’t require synchronization Lower temperature Chasing into final product confirms labeled species is true intermediate Add inhibitor
I1 I1 I2 I2 P P Detecting Intermediates The Challenge: Intermediates are often transient, scarce, and coexist with other molecular species P S I1 I2 Synchronize Reaction To Transiently Enrich Intermediates Pulse-Chase Label Synchronous Cohort Block Reaction Step To Accumulate Intermediate S P S P S I1 I2 I1 I2 I1 I1 P P S { I2 I2 S I2 P S I1 I1 P Partial Reaction P S I1 I2 I2 Examples of blocks: Can provide temporal order of intermediates Can provide temporal order of intermediates Remove or inactivate protein Remove cofactor Label can sensitivity of detection Single molecule analyses follow this principal but don’t require synchronization Lower temperature Chasing into final product confirms labeled species is true intermediate Add inhibitor
I1 I1 I2 I2 P P Detecting Intermediates The Challenge: Intermediates are often transient, scarce, and coexist with other molecular species P S I1 I2 Synchronize Reaction To Transiently Enrich Intermediates Pulse-Chase Label Synchronous Cohort Block Reaction Step To Accumulate Intermediate S P S P S I1 I2 I1 I2 I1 I1 P P S { I2 I2 S I2 P S I1 I1 P Partial Reaction P S I1 I2 I2 Examples of blocks: Can provide temporal order of intermediates Can provide temporal order of intermediates Remove or inactivate protein Remove cofactor Label can sensitivity of detection Single molecule analyses follow this principal but don’t require synchronization Lower temperature Chasing into final product confirms labeled species is true intermediate Add inhibitor
Understanding Molecular Mechanisms S P S = substrate From This: P = product A1 A2 A3 A4 An+1 I = intermediates I4………….. In S I1 I2 I3 P To This: A = activity The transformation of one intermediate into another defines activities which are performed by proteins and/or nucleic acids How does one identify and detect intermediates? How does one structurally characterize intermediates? How does one identify the proteins/nucleic acids responsible for the activities?
Structural Analysis of Intermediates Examples of structural features of intermediates that can be monitored Nucleic Acids Complexes Proteins Composition Cofactor (NTP) Status Size Conformation Stoichiometry Shape Modification Conformation DS versus SS Interacting Sequences Ligand Binding Strand Pairing Interacting Domains Strand Polarity Covalent Linkages Covalent Linkages Modification Topology
Replication is localized to forks (Autoradiograph E. coli DNA) New DNA synthesis is small (pulse label then size) Semi-discontinuous synthesis (EM replicating l DNA) daughter alkaline sucrose gradient EM can distinguish SS from DS DNA small DS DNA (leading) SS DNA (lagging) fork parent HL SS DNA (lagging) DS DNA (leading) fork HL HH daughter (size) Infer H (labeled) and L (unlabeled) from film grain density Detection and Analysis of In Vivo DNA Replication Intermediates SS regions in trans, some interrupted by small DS segments Short (5 - 10 sec) pulse thought to represent initial nucleotide incorporation consistent with discontinuous okazaki fragment synthesis only on strand of specific polarity (lagging)
Replication is localized to forks (Autoradiograph E. coli DNA) New DNA synthesis is small (pulse label then size) Semi-discontinuous synthesis (EM replicating l DNA) daughter alkaline sucrose gradient EM can distinguish SS from DS DNA small DS DNA (leading) SS DNA (lagging) fork parent HL SS DNA (lagging) DS DNA (leading) fork HL HH daughter (size) Infer H (labeled) and L (unlabeled) from film grain density Detection and Analysis of In Vivo DNA Replication Intermediates SS regions in trans, some interrupted by small DS segments Short (5 - 10 sec) pulse thought to represent initial nucleotide incorporation consistent with discontinuous okazaki fragment synthesis only on strand of specific polarity (lagging)
Replication is localized to forks (Autoradiograph E. coli DNA) New DNA synthesis is small (pulse label then size) Semi-discontinuous synthesis (EM replicating l DNA) daughter alkaline sucrose gradient EM can distinguish SS from DS DNA small DS DNA (leading) SS DNA (lagging) fork parent HL SS DNA (lagging) DS DNA (leading) fork HL HH daughter (size) Infer H (labeled) and L (unlabeled) from film grain density Detection and Analysis of In Vivo DNA Replication Intermediates SS regions in trans, some interrupted by small DS segments Short (5 - 10 sec) pulse thought to represent initial nucleotide incorporation consistent with discontinuous okazaki fragment synthesis only on strand of specific polarity (lagging)
DNA Synthesis Occurs Semi-Discontinuously at Replication Forks Leading daughter strand: polymerase moves continuously in same direction as replication fork Lagging daughter strand: polymerase moves discontinuously in opposite direction as replication fork 5’ C 3’ Fork Movement B lagging A 3’ 5’ leading 5’ 3’ Synthesis of Okazaki fragments (1,000-2,000 nt in prokaryotes; 100-200 nt in eukaryote): A. priming B. replacement of primed sequence C. ligation
Replication Fork Tasks and the Activities Activity Task helicase unwind parental strands primase begin DNA synthesis SSBP stabilize SS DNA polymerase synthesize DNA clamp loader/clamp ensure processivity unlink parental strands topoisomerase nuclease/polymerase replace primer ligase connect okazaki fragments
Advantages of Studying Molecular Mechanisms in vitro Can study a process independent of other processes S P S = substrate From This: P = product A1 A2 A3 A4 An+1 I = intermediates I4………….. In S I1 I2 I3 P To This: A = activity The transformation of one intermediate into another defines activities which are performed by proteins and/or nucleic acids How does one identify and detect intermediates? Easier to synchronize or pulse-label the process Easier to block steps or introduce defined intermediates How does one structurally characterize intermediates? Easier to structurally probe or isolate intermediates How does one identify the proteins/nucleic acids responsible for the activities? Can separate and purify activities required for a process
Validating an in vitro system Show the in vitro system shares many properties of the in vivo process Example: replication elongation Substrate DS DNA template; dNTP Product replication fork Intermediates okazaki fragment Genetic Requirements replication mutants Inhibitor Sensitivity aphidicolin (for eukaryotes) Quantitative Properties fork rate okazaki fragment size
Validating an in vitro system Show the in vitro system shares many properties of the in vivo process Example: replication elongation Substrate DS DNA template; dNTP Product replication fork Intermediates okazaki fragment Genetic Requirements replication mutants Inhibitor Sensitivity aphidicolin (for eukaryotes) Quantitative Properties fork rate okazaki fragment size
Phage T4 DNA Replication In Vitro in vivo in vitro 800 nt/sec 500 nt/sec Fork Rate ~ 2 kb ~ 2 kb Okazaki Fragment No OF maturation Genetic Requirements 32, 41, 43, 44, 45, 62 32, 41, 43, 44, 45, 62
Understanding Molecular Mechanisms S P S = substrate From This: P = product A1 A2 A3 A4 An+1 I = intermediates I4………….. In S I1 I2 I3 P To This: A = activity The transformation of one intermediate into another defines activities which are performed by proteins and/or nucleic acids How does one identify and detect intermediates? How does one structurally characterize intermediates? How does one identify the proteins/nucleic acids responsible for the activities?
Purifying Biochemical Activities from In Vitro Systems Fractionation & Reconstitution In vitro complementation Can accelerate by trying to replace fractions with suspected proteins purified in expression systems
Phage T4 DNA Replication In Vitro in vivo in vitro 800 nt/sec 500 nt/sec Fork Rate ~ 2 kb ~ 2 kb Okazaki Fragment No OF maturation Genetic Requirements 32, 41, 43, 44, 45, 62 32, 41, 43, 44, 45, 62 Biochemical activities mostly purified by in vitro complementation Can reconstitute reaction with seven purified proteins
I1 I1 I2 I2 P P Detecting Intermediates The Challenge: Intermediates are often transient, scarce, and coexist with other molecular species P S I1 I2 Synchronize Reaction To Transiently Enrich Intermediates Pulse-Chase Label Synchronous Cohort Block Reaction Step To Accumulate Intermediate S P S P S I1 I2 I1 I2 I1 I1 P P S { I2 I2 S I2 P S I1 I1 P Partial Reaction P S I1 I2 I2 Examples of blocks: Can provide temporal order of intermediates Can provide temporal order of intermediates Remove or inactivate protein Remove cofactor Label can sensitivity of detection Lower temperature Chasing into final product confirms labeled species is true intermediate Add inhibitor
A direct assay for helicase activity * * 41 is NOT required for rapid synthesis on SS DNA 41 has GTP/ATPase activity GTP/ATPase greatly stimulated by SS DNA Inhibition by GTPS slows strand displacement synthesis A Helix Unwinding (Helicase) Activity 41 is required for rapid strand displacement synthesis on DS DNA
Replicative Helicases Many replicative helicases form hexameric rings that are thought to wrap around single-stranded DNA and use the energy of nucleotide hydrolysis to translocate unidirectionally along the DNA, peeling away any hybridized DNA it runs into. These helicases belong to a family of AAA+ ATPases that form multimeric complexes and can couple ATP binding and/or hydrolysis to conformational changes in the complex 5’ 5’ 3’ 3’ 3’ 5’ 5’ 5’ 3’ 3’ 3’ The presumed translocation direction relative to the polarity of the single-stranded DNA defines the polarity of the helicase. Prokaryote: 5’ > 3’ (as modeled above on lagging strand). Eukaryotes: 3’ > 5’ (would be modeled on leading strand).
Okazaki Fragment Maturation E. Coli OF Processing E. coli Phage T4 DNA Pol I (5’ > 3’ Exo) Excise Primer T4 RNaseH Fill-In Gap T4 DNA Pol DNA Pol I Seal Nick T4 DNA Ligase DNA Ligase Addition of T4 RNaseH and T4 DNA Ligase to in vitro replication system produces continuous lagging strand T4 RNaseH or T4 DNA Ligase mutants accumulate okazaki fragments during phage T4 infection in vivo
Replication Fork Tasks and the Activities Activity Task helicase unwind parental strands primase begin DNA synthesis SSBP stabilize SS DNA polymerase synthesize DNA clamp loader/clamp ensure processivity unlink parental strands topoisomerase nuclease/polymerase replace primer ligase connect okazaki fragments
Understanding Molecular Mechanisms S P S = substrate From This: P = product A1 A2 A3 A4 An+1 I = intermediates I4………….. In S I1 I2 I3 P To This: A = activity The transformation of one intermediate into another defines activities which are performed by proteins and/or nucleic acids Other activities may affect the rate, fidelity, specificity, or regulation of these steps How does one identify and detect intermediates? How does one structurally characterize intermediates? How does one identify the proteins/nucleic acids responsible for an activity? How is order, timing, specificity/fidelity, and speed/efficiency maintained?
Processivity How many times an enzyme can act on a substrate before dissociating from it Assay: measure product size under conditions where an enzyme cannot reassociate with its substrate once it dissociates Condition 1: preload enzymes onto substrates then dilute Condition 2: excess substrate (e.g. primer-template) polymerase not processive polymerase processive
An Activity that Enhances Polymerase Processivity 44/62 ATPase and 45 enhance the processivity of T4 DNA polymerase 43 Continuous ATP hydrolysis by 44/62 is not required for enhanced processivity Once ATP is hydrolyzed, processivity factors act like a “sliding clamp” for the polymerase
The Sliding Clamp is a ring that tethers the Polymerase A clamp loader topologically links the clamp to DS DNA Clamp Structure The clamp also interacts with many other proteins to coordinate their function in replication and repair
E. Coli Clamp-Loader (gdd’) loads the Clamp (b ) onto DNA through the ordered execution of activities, each of which is dependent on the substrate generated by the previous activity 3 2 Clamp Loading Model
Summary of Activities and Proteins at the Replication Fork Note: Many of these activities are also required for DNA repair or recombination, and in several cases the same proteins are used Diagram shows prokaryotic 5’>3’ helicase on lagging strand 3’>5’ eukaryotic helicase would be placed on leading strand E. coli Eukaryotes Activity Task DnaB) Mcm2-7, Cdc45, GINS unwind parental strands helicase primase DNA Pol a-primase prime DNA synthesis primase SSBP RPA1-3 stabilize SS DNA SSBP * ** DNA Pol III core DNA Pol e, DNA Pol d synthesize DNA polymerase ensure processivity clamp loader, clamp g-complex, b subunit RFC1-5, PCNA ? coord leading and lagging t subunit Ctf4? unlink parental strands topoisomerase Topo I/Gyrase, Topo IV Topo I/Topo II replace primer polymerase/nuclease DNA Pol I/RNaseH DNA Pol d, FenI, Dna2 connect okazaki fragments ligase DNA Ligase DNA Ligase I * DNA Pol III Holoenzyme ** leading, lagging
Collision Model versus Primer Signaling Model The Challenge of Regulating and Coordinating Multiple Activities Primase synthesizes primer Clamp-loader positions clamp around primer-template Polymerase loads onto primer-template and binds to clamp Primase synthesizes primer for next okazaki fragment Polymerase synthesizes okazaki fragment What regulates primer synthesis? Polymerase dissociates from clamp to load onto next primer Clamp-loader loads clamp What regulates polymerase processivity? Okazaki fragment maturation is completed Clamp-loader eventually releases clamp for reuse on other okazaki fragments What directs when clamps are released? Adapted from Molecular Biology of the Cell. 4th Ed.
Support for Collision Model of Lagging Strand Polymerase release Pol III* - clamp association is sensitive to Pol III colliding into 5’ end all 4 dNTPs Mimic “replicating” Pol III by idling with only 2 dNTPs Pol III completes replication and runs into back of primer Pol III* stably associates with clamp on DNA Pol III* rapidly dissociates from clamp on DNA and within 1 sec reassociates with clamp preloaded on new primer-template
Keeping the Lagging Strand Polymerase at the Replication Fork Processive synthesis of okazaki fragments by lagging strand polymerase suggests tethering to leading strand replication proteins at the fork, generating a dynamic lagging strand loop (trombone model). In E. coli, tau dimer tethers by binding two core polymerases in the Pol III holoenzyme B clamp core • Complex clamp-loader t dimer core B clamp Pol III holoenzyme Predicted lagging strand “loop” seen in EM; dynamic loop behavior detected by single molecule analysis Figures from Molecular Biology of the Cell. 4th Ed.
Trombone Model from Cell Snapshots (Cell 141:1088) How many polymerases can interact with each clamp? How do primase and helicase interact yet work in opposite directions? What holds leading and lagging strand polymerases together in other systems? Are leading and lagging polymerization coordinated? See Movie on Bioreg Website Links Page
5’ 5’ 5’ 5’ 5’ 5’ 5’ 5’ 5’ 5’ Lecture 1: DNA Polymerase Use of biochemistry (assays) and genetics (phenotypes) to define function Structure critically informs but does not define function Fidelity/Specificity: bioregulation through substrate control of molecular choice The Replisome Lecture 2: Breaking down complex processes into intermediates and subreactions In vivo and in vitro analysis of the players, intermediates, and activities Defining activity dependencies to understand their order and timing
General Strategies for Isolating DNA Synthesis Mutants 1) Screen: isolate ts-lethal mutants then assay macromolecular synthesis soon after shift to restrictive temp • DNA synthesis– (3H Thymine incorp.) • RNA synthesis + • Protein synthesis + • Nucleotide synthesis + • 2) Selective enrichment: ts mutants that fail to incorporate poisonous nucleotide analog during transient shift to restrictive temp wash out 5-BU 5-BU 32° 42° 42° 32° UV WT incorporate 5-bromouracil (5-BU) - UV sensitive Replication mutants poorly incorporate 5-BU - survive
Functional analysis of replication mutants limited by in vivo assays generation time Log DNA synthesis Time shift to restrictive temperature Slow Stop: involved in initiation ongoing replication continues new rounds blocked Quick Stop: involved in elongation ongoing replication blocked
Assigning an activity to a protein(s)/nucleic acid(s) What is necessary for an activity? (can be asked in vitro and in vivo) Does absence or inactivation of a protein/nucleic acid lead to accumulation and disappearance of the appropriate intermediates? What is sufficient for an activity? (best asked in vitro) Can the key feature of the activity be assayed in isolation? Can a purified protein(s) and/or nucleic acid(s) perform that isolated activity?
Polymerase/primase Coupled Assay 61 Direct Analysis by TLC, PAGE, or RNA sequencing An Activity that Primes DNA Synthesis (Primase) 41 (helicase) and 61 (primase) can act together as a mobile primosome that has much better helicase and primase activity than either alone.
Edwards lab, CSHSQB (2000) 65:193-200 SS DNA Binding Protein helps present and protect SS DNA Binds cooperatively to single-stranded DNA and stretches out the phosphate backbone protein-DNA and protein-protein binding energy stabilizes unwound DNA and makes unwinding energetically less costly prevents secondary structure from interfering with enzyme function Protects sugar-phosphate backbone from nucleases while leaving bases available for pairing RPA (eukaryotic SSBP) bound to 27 nt SS DNA side view down axis of SS DNA { bases accessible
Topoisomerases Solve Topological Problems During Replication Daughter DNA can be left catenated after termination Fork movement can generate superhelical tension converging replication forks Two fundamental classes of topoisomerases: Type I can transiently break single-stranded DNA and pass another single-strand through the break Type II can transiently break double-stranded DNA and pass another double-strand through the break Each enzyme class holds onto the broken DNA ends by reversible covalent linkage Although not intuitively obvious, both strand passing events will relieve superhelical tension Only Type II topoisomerases, however, can decatenate DS DNA Type II mechanism Type I mechanism Cross-section of passed strand Figures from Wang Lab, Nat Rev Mol Cell Biol. (2002) 3:430
Advantages of In Vitro Systems for Mechanistic Studies A1 A2 A3 A4 An+1 I4………….. In S I1 I2 I3 P Isolating the Process: Can study a process independent of other processes Identifying the Players: Can purify activities required for a process Temporal/Kinetic Analysis: Easier to synchronize or pulse-label the process Establishing Partial Reactions: Easier to block steps or use defined intermediates Characterizing Intermediates: Easier to probe or isolate intermediates






















