Download
1 / 1

E N D
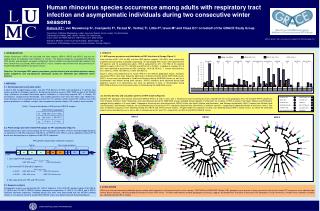
HEV: 6 (3%) 1. INTRODUCTION Human rhinoviruses (HRVs) are classified into three species: HRV-A, HRV-B and HRV-C and are the leading cause of respiratory tract infections in humans. The disease outcomes associated with different HRV species and genotypes are poorly established. Several studies have documented high incidence and more severe illness caused by HRV-A and HRV-C viruses, however most of them have focused mainly on hospitalized pediatric patients during one season. Objective: To investigate HRV species prevalence, genetic diversity and circulation patterns among adults outpatients and asymptomatic individuals during the 2007/2008 and 2008/2009 winter seasons. ANTWERP BE GENT BE UTRECHT NL ROTENBURG DE CARDIFF UK SOUTHAMPTON UK BARCELONA SP MATARO SP LODZ PL SZCZECIN PL BIALYSTOCK PL 20 18 16 14 12 Number of genotyped HRVs 10 8 6 4 2. METHODS 2.1. Clinical specimens and study cohort A total of 482 nasopharyngeal swabs, real-time PCR positive for HRV, were collected at 11 primary care centers located in 6 European countries, during two consecutive seasons (2007-2009) as part of the GRACE study (Genomics to combat Resistance against Antibiotics in Community-acquired LRTI in Europe, EU framework 6, funded contract PL518226). The studied specimens were from 481 adults (age 18-89 years; mean 48 years) with a respiratory disease at their first (V1 samples) and/or second (V2 samples) visit to the general practitioner. In addition, samples from asymptomatic control subjects (V0 samples) were available. 2 0 Epidemic seasons 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 07/08 08/09 N.S.* Genotyped HRVs (%) 29% 68% 25% 52% 44% 17% 45% 44% 36% 4% 38% 71% 24% 70% 44% 48% 75% 44% 21% N.S.* 63% HRV-A: 131 (67%) HRV-B: 26 (13%) HRV-C: 33 (17%) N.S.*: no samples available for this season ATWERP (BE) BARCELONA (SP) GENT (BE) MATARO (SP) UTRECHT(NL) LODZ (PL) 2007/2008 2008/2009 ROTENBURG (DE) BIALYSTOCK (PL) 148 (76%) 217 (76%) V1 CARDIFF (GB) SZCZECIN (PL) Table 1. Seasonal distribution of HRV positive GRACE samples SOUTHAMPTON (GB) GENBANK 43 (15%) 30 (15%) V2 27 (9%) 17 (9%) V0 V1(2)*-07/08 V1-08/09 V1-08/09 V1-08/09 HRV-A HRV-B HRV-C HRV32 V1-08/09 HRV67 HRV9 V1-08/09 V1-08/09 HRV95 V1-08/09 HRV8 HRV45 V2-07/08 HRV60 HRV38 V1-08/09 V0-08/09 V1-08/09 V1-08/09 V1-08/09 HRV94 V1-07/08 HRV70 100 V1-08/09 V1-08/09 HRV64 V1-08/09 HRV74 100 HRV19 V1-07/08 HRV15 V1-08/09 V2-07/08 V1-07/08 AU-QPM/2003 V2-08/09 HRV82 100 0.05 2.2. Primer design and semi nested PCR assays for VP1 amplification (Figure1) Degenerate primers were used to amplify the VP1 capsid gene of HRVs. Primers were developed based on an alignment of 139 HRV sequences (98 HRV-A, 32 HRV-B and 9 HRV-C) and an additional internal VP1-B primer was designed from an alignment of 84 HRV-B sequences. HRV17 HRV53 V1-08/09 V1-08/09 HRV20 0.05 V1-V2*-07/08 HRV28 V2-08/09 100 97 100 V1-08/09 100 HRV91 V1-08/09 99 HRV68 V1-08/09 100 99 87 V1-08/09 100 V1-08/09 0.05 V0-07/08 HRV69 100 V1-08/09 V1-08/09 100 99 100 HRV71 99 HRV84 HRV96 100 81 A101 100 100 100 100 HRV61 HRV51 HRV73 100 100 100 HRV13 HRV65 100 HK-026/2005 HRV41 V1-07/08 100 V1-08/09 99 HRV97 V1-07/08 99 V1-08/09 95 100 99 100 100 V1-08/09 V0-07/08 100 100 HRV80 V1-08/09 100 100 92 100 HRV23 HRV46 100 97 100 V2-07/08 V1-07/08 92 HRV genome organization and primers locations HRV78 V1-08/09 100 82 HRV93 HRV7 99 HRV30 97 V0-07/08 99 92 99 100 96 100 HRV88 V1-08/09 99 100 V1-07/08 100 100 100 HRV49 100 HRV58 HRV52 100 100 Capsid Non-structural proteins 100 HRV89 HRV27 HRV2 CA-NAT001/2001 79 100 100 HRV36 V1-08/09 94 100 96 96 100 V0-07/08 99 V1-08/09 100 100 V0-07/08 V1-08/09 P1 P2 P3 87 V1-07/08 87 V1-07/08 V2-07/08 Shanghai-N4/2006 100 V1-07/08 100 V1-08/09 V1-08/09 V1-08/09 100 99 V1-08/09 V1-08/09 HRV62 V1-08/09 97 3B 3Cpro 3Dpol 99 V4 V2 V3 V1 2A 2B 2C 3A V1-08/09 100 HRV25 HRV26 100 100 V1-07/08 V1-07/08 100 5’-UTR 3’-UTR 100 100 100 V1-08/09 V1-08/09 V1-08/09 100 100 HRV48 V1-08/09 99 100 99 96 100 98 V1-08/09 83 100 V1-08/09 83 99 V1-07/08 100 V1-08/09 HRV99 92 100 1. One-step RT-PCR (Qiagen) 98 NY-074/2004-2005 99 100 100 V1-08/09 82 V1-08/09 96 90 HRV43 V1-08/09 HRV47 HRV75 HRV31 100 V1-08/09 100 99 77 94 76 100 V1-08/09 100 V1-08/09 V1-08/09 ~1.5 kb HRV42 V1-08/09 HRV-VP3 sense HRV-2A antisense 99 100 V1-07/08 HRV34 100 100 100 100 V1-08/09 V1-08/09 99 100 HRV50 100 V1-07/08 HRV29 97 HRV18 100 100 100 100 HRV44 HRV5 100 V1-08/09 100 V1-07/08 V2-08/09 2. Semi-nested PCR (Biorad IQ supermix) 100 100 100 V1-08/09 100 100 100 V1-07/08 100 99 V1-08/09 100 100 V1-08/09 HRV83 V1-08/09 Shanghai-N10/2007 98 100 HRV77 V1-08/09 100 V1-08/09 HRV100 V1-08/09 HRV10 HRV86 HRV21 100 V1-08/09 V1-07/08 V1-08/09 V1-08/09 100 ~1 kb V2-08/09 HRV57 V1-08/09 sn-PCR1 HRV-VP3 sense HRV-VP1 antisense HRV39 HRV92 HRV63 V2-08/09 V2-07/08 V1-08/09 V0 -07/08 V1-08/09 100 100 HRV59 HRV6 V2-08/09 HRV33 HRV40 HRV11 V1-08/09 HRV76 V2-07/08 V1-08/09 HRV90 HRV-VP1-B antisense sn-PCR2 HRV-VP3 sense HRV85 HRV24 HRV79 HRV56 V1-07/08 HRV1B CA-NAT045/2003 V1-07/08 V1-08/09 HRV1 V1-07/08 V1-08/09 V1-08/09 HRV37 V1-08/09 HRV16 HRV98 HRV54 HRV3 V2-07/08 HRV35 HRV81 V2-07/08 V1-08/09 V1-08/09 V1-08/09 V1-07/08 HRV72 HK-024/2005 HRV14 3. ABI sequencing with VP3 and VP1 primers HK-025/2005 2.3. Sequence analysis Phylogenetic analysis was performed with ~800 nt fragments of the VP3-VP1 genome region of 90 HRV-A, 17 HRV-B and 19 HRV-C GRACE isolates compared to respectively 71 HRV-A, 24 HRV-B and 9 HRV-C GenBank reference sequences. Neighbor-joining (NJ) trees were constructed with the MEGA4 software (Figure 3). The genetic similarity among HRV GRACE strains was determined with GeneDoc program. Human rhinovirus species occurrence among adults with respiratory tract infection and asymptomatic individuals during two consecutive winter seasons Zlateva KT1, van Nieuwkoop S1, Coenjaerts F2, Farzad N1, Verheij T3, Little P4, Ieven M5 and Claas EC1 on behalf of the GRACE Study Group 1Department of Medical Microbiology, Leiden University Medical Center, Leiden, The Netherlands 2Deparment of Virology, UMC Utrecht, Utrecht, The Netherlands 3Julius Center for Health Sciences and Primary Care, UMC Utrecht 4School of Medicine, University of Southampton, Southampton, UK 5Department of Microbiology, University Hospital Antwerp, Antwerp, Belgium GRACE project: https://www.grace-lrti.org/portal/en-GB/Homepage.htm 3. RESULTS 3.1. HRV species prevalence and distribution of HRV infections in Europe (Figure 2) From the total of 357 (74% of 482) real-time HRV positive samples, 196 (55%) were successfully amplified and analysed by nucleotide sequencing. The genotyped HRV strains were obtained from 179 patients and 16 asymptomatic adults. HRV-A accounted for 67% (131/196) of the positive GRACE samples, HRV-C for 17% (33/354) and HRV-B 13% (26/357). In addition 6 samples (3% of 357) were identified to be positive for HEV, including 1 HEV 68 (HEV-D), 1 Human coxsackievirus A13 (HEV-C), 2 HEV 104 (HEV-C), 2 HEV 109 (HEV-C). Figure 2 shows the predominance of mainly HRV-A in the different geographic origins. Infections caused by HRV-C were more frequently observed in Antwerp and Gent during 2007/2008 season. HRV-C viruses were not detected in Cardiff and Szczecin during 2008/2009 season as well as in Southampton and Mataro during both seasons. HRV-B infections were observed during both epidemic seasons in 6 communities, but were not detected in Gent, in Southampton during 2007/2008 and also in the three communities in Poland during the 2008/2009 season. 3.2. Genetic diversity and circulation patterns of HRV isolates (Figure 3) Multiple VP1 genotypes were present among the GRACE HRV-A, B and C strains with a tendency for close clustering of strains isolated from the same geographic area. For example HRV-A sequences from Antwerp, Szczecin, Gent, Rotenburg, and Lodz obtained during the 2008/2009 season, grouped closely together. Furthermore the majority of HRV-A isolates from Spain (Mataro and Barcelona) grouped closely together in 3 main clades. Geographic clustering was also observed for HRV-B strains from Spain (Mataro and Barcelona), and Antwerp and between HRV-C isolates from Poland (Lodz and Bialystock), and Antwerp. However, we also detected close clustering of VP1 sequences obtained from communities in different countries, suggesting for global distribution of rhinovirus strains. Two pairs of identical sequences were identified among the HRV-B and HRV-A clinical isolates. A higher degree of sequence variation was detected among HRV-C clinical strains, for which theestimatedpaiwiseamino acididentity was as low as 58% in comparison to 63% and 72%, respectively, for HRV-A and HRV-B GRACE viruses. 4. CONCLUSIONS HRV-A was the most commonly detected species among adult outpatients in Europe during the winter seasons 2007/2008 and 2008/2009. Multiple HRV genotypes were present in each community and closely related VP1 sequences were obtained from several distant locations, indicating broad distribution of certain HRV strains. The observed tendency towards geographic clustering, suggests for predominant circulation of particular HRV genotypes in each community. A higher level of genetic variability was identified among HRV-C isolates. Leiden University, Leiden, The Netherlands












