Download
1 / 48
540 likes | 945 Views
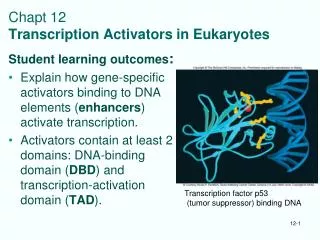
Chapt 12 Transcription Activators in Eukaryotes. Student learning outcomes : Explain how gene-specific activators binding to DNA elements ( enhancers ) activate transcription. Activators contain at least 2 domains: DNA-binding domain ( DBD ) and transcription-activation domain ( TAD ).

E N D
Chapt 12Transcription Activators in Eukaryotes Student learning outcomes: • Explain how gene-specific activators binding to DNA elements (enhancers) activate transcription. • Activators contain at least 2 domains: DNA-binding domain (DBD) and transcription-activation domain (TAD). Transcription factor p53 (tumor suppressor) binding DNA
Describe some DBD and TAD motifs: • Zinc modules, • homeodomain, bZIP, bHLH (shared with prokaryotes) • Examples of activators • Explain how activators function by contacting GTFs, recruiting pol II or contacting coactivators • Describe how activators and coactivators can be controlled by modifications including: ubiquityation, sumoylation, phosphorylation, acetylation • Important Figures: 1, 2*, 3, 4*, 6, 9, 10*, 12*,16, 18*, 20, 23, 24, 27, 33*, 34*, 35, 36, 37; Table 1 • Review questions: 1, 2, 3, 4, 5, 6, 9, 10, 13, 14, 17*, 18, 24, 28, 30, 31; Analy Q 1
12.1 Categories of Activators • Activators can stimulate or inhibit transcription by RNA polymerase II • Composed of at least 2 functional domains: • DNA-binding domain (DBD) • Transcription-activation domain (TAD) • Many also have dimerization domain • Domain - independently folded region of protein Ex. Fig. 13 Gal4 activator of yeast
DNA-Binding Domains (DBD) DNA-binding domains - DNA-binding motifs: • 3 Characteristic shapes for specific DNA binding Zinc-containing modules: use Zinc ions to create shape to fit a-helix of motif into DNA major groove • Zn fingers, Zn modules, Modules with 2 Zn and 6 cys Homeodomains: about 60 amino acids - resemble HTH (helix-turn-helix) proteins in structure and function; ex. homeobox proteins regulate fruit fly development bZIP and bHLH motifs: highly basic DNA-binding motif linked to protein dimerization motifs • Leucine zippers, Helix-loop-helix (ex. CCAAT enhancer-binding protein)
Transcription-Activating Domains (TAD) Most activators have at least one of these domains: • Acidic domains – ex. yeast GAL4 (11 acidic amino acids out of 49 amino acids) in domain • Glutamine-rich domains – ex. Sp1 has 2 regions that are 25% glutamine (Gln) • Proline-rich domains – ex. CTF has domain with 19 proline in 84 amino acids
12.2 Structures of DNA-Binding Motifs of Activators • DNA-binding domains well-defined structures (Ch. 9) • X-ray crystallography shows how structures interact with DNA targets • Interaction domains often form dimers, or tetramers, • Most DNA-binding proteins can’t bind DNA in monomer form • Compare common DNA binding motif in prokaryotes
Helix-Turn-Helix motif is common:lac, trp, l, other phage repressors l Family of Repressors Repressors have recognition helices (red) fit major groove of operator • Helix-turn-helix motif (HTH) • Specificity of binding depends on amino acids in recognition helices Fig. 9.2
High-Resolution Analysis of l Repressor-Operator Interactions Structural Features from co-crystal: • Recognition helices of repressor monomer (3, red) nestle into DNA major grooves in 2 half-sites • Helices approach, hold two monomers together in repressor dimer • DNA similar in shape to B-form DNA • DNA bends at two ends of fragment as it curves around repressor dimer Figs. 9.6, 9.7: Repressor dimer binding to symmetric operator O1
Eukaryotic Homeodomains have HTH motif • Homeodomains contain DNA-binding HTH (helix-turn-helix) motif • Recognition helix (3) fits in DNA major groove, makes specific contacts • N-terminal arm nestles in adjacent minor groove • First identified Drosophila developmental genes – Fig. 9 Drosophila homeotic engrailed protein binding target Fig. 8 Antennapedia mutant has legs in place of antennae
Eukaryotic bZIP and bHLH Domains • bZIP proteins dimerize through leucine zipper motif • Leu every 7 aa (one face); hydrophobic interactions • adjacent basic regions embrace DNA like tongs • bHLH proteins dimerize through helix-loop-helix motif • basic parts of each long helix grasp DNA target site • Ex. bZIP yeast GCN4 activator Fig. 10 structure leucine zipper Fig. 11 bZIP of yeast GCN4 activator
Eukaryotic bZIP and bHLH Domains • bHLH motif in activator myoD: • HLH is dimerization • Long helix 1 has basic region that binds DNA • Both bHLH and bZIP domains are found in Myc, Max oncogenes; heterodimers bind to DNA like HLH; extra dimerization potential from leu zippers
Zinc Fingers Fig. 1 3D shape of 1 Xenopus Zn finger: Zn blue; Cys yellow S; His blue • First described forTFIIIA • 9 repeats of 30-aa element: • 2 closely spaced Cys followed 12 aa by 2 closely spaced His • Coordination of aa to metal helps form finger-shape • Protein rich in Zn: 1 Zn per repeat • Specific recognition between Zn finger and DNA in major groove Fig. 2; Zn finger has antiparallel b-strand (2 Cys) and a-helix (2 His)
3 Zinc Fingers in Curved Shape bind DNA Zif268 is mouse TFIIIA-class protein Each Zn finger: • antiparallel b-strand contains 2 Cysteines • 2 Histidines in a-helix • Helix and strand coordinated to Zn ion aa of helix recognize bp of major groove Fig. 3
Zn module: GAL4 Protein of Yeast • GAL4 regulates expression of genes for galactose metabolism • Binds target UAS (upstream activating sequence) • Member of Zn-containing family of DNA-binding proteins, Not zinc finger • Each GAL4 monomer contains DNA-binding motif: • 6 Cys coordinate 2 Zn ions in a bimetal thiolate cluster • DNA recognition - short a-helix protrudes into major groove • Dimerization motif on other end - a-helix forms parallel coiled coil, interacts with a-helix on other GAL4 monomer
Zn module - GAL4 protein of yeast Fig. 4 dimer of GAL4 binding DNA: Zn yellow DNA recognition aa 8 to 40; linker 41-49; dimerization 50-64
Zn modules - Nuclear Receptors • Nuclear hormone receptors interact with different endocrine-signaling molecules • Protein plus endocrine molecule forms complex that functions as activator: • Binds hormone response elements, stimulates transcription of associated genes • Ligands for these receptors are lipid-soluble, cross membrane (steroids, retinoic acid, vitamin D)
Type I Nuclear Receptors • Type 1 include Glucocorticoid (GR), Estrogen (ER) • Reside in cytoplasm bound to protein Hsp90 • When receptors bind to hormone ligands: • Release cytoplasmic protein partners • Move to nucleus; dimerize • Bind to enhancers • Act as activators Fig. 6
Glucocorticoid Receptor binding DNA • DNA-binding domain with 2 Zn-containing modules • One module has most of DNA-binding residues • Other module has surface for protein-protein interaction to form dimers Fig. 6 structure with half sites separated by extra bp; only 1 module binds best
Types II and III Nuclear Receptors • Type II nuclear receptors stay within nucleus • Examples: thyroid hormone receptor (TR), • vitamin D receptor (D3) • Retinoic acid receptor (RAR) (more Ch. 13) • Bound to target DNA sites • Without ligands, receptors repress gene activity • When receptors bind ligands, activate transcription • Type III receptors are “orphan” whose ligands are not yet identified • All use a-helix for interaction with DNA sequences
12.3 Independence of Domains of Activators • DNA-binding, transcription-activating domains of activator proteins are independent modules • Hybrid proteins with DNA-binding domain of one protein, transcription-activating domain of another • Hybrid protein still functions as activator Fig. 18; yeast expressGal4 TAD on Gal4 or lexA DBD
12.4 Functions of Activators for pol II • Eukaryotic activators recruit pol II to promoters • Stimulate binding of general transcription factors (GTFs) and pol II to promoter • 2 hypotheses for recruitment: • GTF cause stepwise build-up of preinitiation complex • GTF and other proteins already bound to polymerase in complex called RNAP holoenzyme Kornberg lab quantitation of proteins tested models
Models for Recruitment Fig. 14
Recruitment Model of GAL11P-containing Holoenzyme • Ptashne found dimerization domain of GAL4 binds to mutant form of Gal 11 (GAL11P) in holoenzyme (GAL11P is potentiator) [GAL11 is partof mediator complex] • Holoenzyme, plus TFIID, then binds to promoter, activating gene Fig. 16
Recruitment Model of GAL11P-containing Holoenzyme • Activation in some yeast promoters appears to function by recruitment of holoenzyme • chimeric activator proteins with lexA DBD and other domains recruit holoenzyme Fig. 17
Recruitment of holoenzyme may not be that common • Kornberg evidence: • TAP- epitope tags on components: • Antibodies on immunoblot quantified pol II, GTFs, mediator: • pol II and some GTFs present in much higher concentration • Unlikely holoenzyme recruited as unit Fig. 18 Pol II about 3x more than Mediator, TFIIH
12.5 Interaction Among Activators • General transcription factors (GTFs) interact to form preinitiation complex (PIC) • Activators and GTFs also interact • Activators interact with one another to activate gene • Individual factors interact to form protein dimer facilitating binding to single DNA target • Specific factors bound to different DNA targets can collaborate to activate gene
Importance of Dimerization to Activation • Dimerization increases affinity between activator and DNA target: Some activators form homodimers (ex. GAL4) Some form heterodimers Ex. Products of jun and fos Oncogenes form heterodimer, bind AP-1 sites; heterodimer Binds better than homodimers Fig. 19; EMSA assay; Fos or Fos Core mixed with Jun or Jun core and AP-1 site; M = myc oncoprotein
Activators can Act at a Distance • Bacterial and eukaryotic enhancers stimulate transcription even though located some distance from promoters • Hypotheses explain ability of enhancers to act at a distance • Change in topology • Sliding • Looping ** • Facilitated tracking Looping brings activators bound to different enhancers closer to promoter
Hypotheses of Enhancer Action Fig. 20: a. Topology b. Sliding c. Looping d. Tracking
Complex Enhancers • Many genes have more than one activator-binding site, -> respond to multiple stimuli • Each activator that binds must interact with PIC assembling at promoter, likely by looping out intervening DNA • Consider whole region upstream of promoter as an enhancer (rather than each specific site for each factor); different factors binding permit fine level of control on expression • Combinatorial code: final result of all activators, inhibitors bound
Complex enhancer: Control Region of Metallothionein Gene Fig. 23 • Gene product helps eukaryotes cope with heavy metal poisoning (small proteins rich in cys; bind metals) • Turned on by several different agents BLE = basal level enhancer; MRE = metal response element; GRE = glucocorticoid response element; GC = Sp1 binding
Complex enhancers in development Figs. 24, 25: sea urchin Endo16 gene: different activators and modules at different times control expression Red = enhancers; Blue = architectural factors Ovals - activators
Architectural Transcription Factors Architectural transcription factors: main purpose to change shape of DNA control region so other proteins bind successfully to stimulate transcription HMG proteins bend DNA, help activators bind (HMG, small nuclear proteins with high electrophoretic mobility) ex. LEF-1 (T-cell receptor); HMGI (Y) for interferon B gene
Architectural Transcription Factor • T cell receptor a-chain: enhancer elements upstream • Elements bind to: Ets-1 LEF-1 CREB • CREB (cyclic AMP response element binding protein) • LEF-1 (lymphoid enhancer) not enhance transcription alone • Activator LEF-1 binds minor groove of DNA through HMG domain, induces strong bending of DNA • DNA bending helps other activators bind, and interact with activators and general transcription factors Fig. 26 T cell receptor a chain
Enhanceosome • Complex of enhancer DNA with activators contacting DNA • Ex. HMG helps bend DNA so that it interacts with other proteins Fig. 27 • HMG I(Y) plays similar role in human interferon-b control gene: • Activation seems to require cooperative binding of several activators, including HMG I(Y) to form enhanceosome with a specific shape
Insulators prevent activation of unrelated genes by nearby enhancers: • Enhancer-blocking activity: insulator between promoter and enhancer prevents promoter activated • Barrier activity: insulator between promoter and condensed, repressive chromatin prevents promoter repressed Fig. 28
Mechanism of Insulator Activity • Sliding model • Activator bound to enhancer and stimulator slides along DNA to promoter • Looping model • Two insulators flank an enhancer; when bound, interact to isolate enhancer Fig. 29
12.6 Regulation of Transcription Factors • Hormones binding nuclear receptors caused them to move to nucleus, or to change from repressor to activator (Ch. 13) • Phosphorylation of activators allows to interact with coactivators to stimulate transcription • Ubiquitylation of transcription factors marks them for • Destruction by proteolysis • Stimulation of activity • Sumoylation - attachment of polypeptide SUMO; targets movement to compartments of nucleus • Methylation and acetylation can modulate activity
Coactivators mediate signals from activator to basal transcription apparatus • CBP/p300 is common coactivator: CREB binding protein/ 300kD protein HAT activity, acetylates histones and others • CREB (cyclic AMP responsive element binding protein) binds CRE (Fig. 33) • Nuclear receptors plus hormone bind DNA (Fig. 34) • Both effects mediated through CBP
Coactivators do not stimulate basal transcription, only activated • Mediator: (Kornberg) • Mediates activation; binds CTD of pol II • Found in yeast first • Many similar complexes: • SMCC/ TRAP • CBP/p300 (CREB-binding) • CRSP (for SP1) • SRC (steroid receptor co-) Fig. 32
Coactivators Mediate Activation Ex. CREB stimulates basal transcription machinery via CBP/p300 Protein Kinase A functions insignal transduction pathway Phosphorylation of CREB by PKA is critical for activation; Fig. 33
Activation of Nuclear Receptor-Activated Gene uses CBP/ SRC coactivator CARM1 methylates proteins Fig. 34
Ubiquitylation affects activation • Ubiquitin - 76-aa polypeptide covalently attached to Lys residues by Ubiquitin ligase • Ubiquitylation, especially monoubiquitylation, of activators can have activating effect • Polyubiquitylation marks proteins for destruction in proteasome, reduces or abolishes activation • Proteins from 19S regulatory particle of proteasome can stimulate transcription
Sumoylation affects Activation • Sumoylation - addition of one or more copies of 101aa polypeptide SUMO (Small Ubiquitin-Related Modifier) to Lysine residues; similar to ubiquitylation • Sumoylated activators targeted to a nuclear compartment that keeps them stable, but not activating targets (PML nuclear bodies) • Ex. PML protein in • promyelocytic leukemia; • B-catenin and LEF-1 SUMO Some proteins in nuclear bodies
Acetylation affects Activators • HAT - enzyme histone acetyltransferase, modifies histone tails with acetyl groups (Ch. 13) • Activity of CBP/p300 • Nonhistone activators and repressors can be acetylated by HATs • Acetylation has either positive or negative effects
Signal Transduction Pathways • Begins with signaling molecule interacting with receptor on cell surface (hormones, growth factors) • Interaction sends signal into cell (second messenger pathway) • Often leads to altered gene expression • Often protein phosphorylation passes signal from one protein to another (PKA, MAPK, G proteins) • Signal amplification occurs at each step
Three Signal Transduction Pathways that share coactivator CBP/p300
Review questions • 1. List three different classes of DNA-binding domains found in eukaryotic transcription factors. • 2. List three different classes of transcription-activation domains in eukaryotic transcription factors. • 7. Explain the difference between type I and II nuclear receptors, and give an example of each. • 28. Draw diagrams to illustrate action of CBP as activator of (a) phosphorylated CREB; (b) nuclear receptor