Download

1 / 33
330 likes | 518 Views
1. Fibroblast Chemotaxis: more about positive feedback loops. 2. Autoregulatory Mechanisms of Eukaryotic Chemotaxis System Components: Receptors, G-proteins, GEFs, PI3K, Kinases, phosphatases. How evolution has selected for components with autoregulation and integral feedback control.

E N D
1. Fibroblast Chemotaxis: more about positive feedback loops. 2. Autoregulatory Mechanisms of Eukaryotic Chemotaxis System Components: Receptors, G-proteins, GEFs, PI3K, Kinases, phosphatases. How evolution has selected for components with autoregulation and integral feedback control.
Fibroblasts chemotax toward growth factors 0 3 hrs 8 hrs 12 hrs 21 hrs PDGF-stimulated wound healing in mouse embryo fibroblasts
Regulation Tissue Tyr Kinase All Tyr Kinase + b/g All Tyr Kinase Blood Cells b/g Blood Cells PI3K p110 Family Members Class Ia 110 a p 8 5 R a s Kinase C2 PIK b i n d i n g b i n d i n g 110 b 110 d Class Ib 110 g
Deletion of Class Ia PI3K genes in mouse embryo fibroblasts impairs PDGF-dependent cell migration. Brachmann et al., 2005 Mol. Cell. Biol. 25, 2593. 0 3 hrs 8 hrs 12 hrs 21 hrs WT PI3K-Ia Deletion
Woundhealing 10ng/ml PDGF 80 3h 8h 15h 21h 70 60 Migrated Cells 50 Ly294003 PI3K inhibitor 40 30 20 10 0 Wild type P85a-/-;p85b-/- PI3K Ia deletion
Defect in PDGF-induced lamellipodia formation in MEFs defective in class Ia PI3K Brachmann et al., 2005 Mol Cell Biol 25, 2593 unstimulated PDGF PDGF + WM Wild Type PI3K Ia deletion
GPCR bg p110b can also be activated by bg subunits of G proteins, but only when bound to a phosphoTyr protein (AND GATE). Class Ia PI3K has multiple domains for signal input, allowing it to act as an ‘AND GATE’or possibly an ‘OR GATE’ Membrane GTP Ras GTP CDC42 Tyr Kinase P-Tyr P-Tyr SH3 Cdc42 binding SH2 SH2 Ras Binding PIK Catalytic C2 p85 regulatory p110 catalytic Class Ia PI 3-Kinase
PIP3 PIP3 PTEN PH GTP Rac p110 PI3K p85 SH2 AKT Cortical Actin Cell Migration Class Ia PI3K mediates growth factor-dependent cortical actin formation Growth Factor Receptor PIP2 Ras Tyr Kinase GTP Rac GEF? P-Tyr
Deletion of class Ia PI3K genes appears to impair (but not eliminate) Ras activation (as judged by impaired activation of the downstream protein kinase, Erk) Brachmann et al., 2005 Mol Cell Biol. 25, 2593 5 min PDGF [ng/ml]: 0 1 3 10 0 1 3 10 Erk-P Erk Wild Type MEFs PI3K Ia deleted Thus, as in Dictyostelium, there appears to be a positive feedback loop between PI3K and Ras in fibroblasts.
Reduced PDGF-induced Rac activation in MEFs lacking class Ia PI3K Brachmann et al., 2005 Mol Cell Biol. 25, 2593 Control Double KO PDGF: - + - + Rac GTP GST-CRIB pulldown Rac p85a Erk-P Erk
Overexpression of a Rac GEF (Vav2) induces lamellipodia formation in MEFs lacking Class Ia PI3K Brachmann et al., 2005 Mol Cell Biol. 25, 2593 Wild Type PI3K Ia deleted Rhodamine-Phalloidin (Actin) Vav2
PI3K is involved in both local Ras and local Rac positive feedback loops ? Ras GTP PDGF Receptor p110 p85 PIP3 + PH GEF Rac GTP ? +
Conclusions 1. Growth Factor Receptors stimulate Class Ia PI3K through PhosphoTyr residues of receptors binding to SH2 domains, while GPCRs stimulate Class Ib PI3K through bg subunits binding to the catalytic subunit. 2. In both cases, PI-3,4,5-P3 is in a local positive feedback amplification loop involving Rac (and Ras?) that allows non-isotrophic localization of cortical actin, providing directionality to chemotaxis.
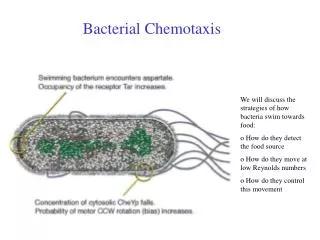
How is perfect adaptation achieved in eukaryotic chemotaxis? Shutoff mechanisms must exist to adapt the system to a given level of stimulation, allowing a temporal increase in receptor stimulation to be sensed. The adaptation should be slow compared to the stimulation to insure significant directional migration prior to adaptation. What is known about shutoff mechanisms of GPCRs and Receptor Tyr Kinases?
GPCR ACTIVATION Hormone Receptor a GDP bg Receptor a GDP bg
GPCR ACTIVATION attractant Receptor Receptor Receptor a a GTP bg bg bg Effector 2 GDP GTP PI 3-kinase etc. a GTP Effector 1 (Phospholipase C, etc.) Ligand-induced Conformational Change <100 msec msec to sec Receptor a GDP bg
Signal Termination, Downregulation and Reset to Basal State RGS Seconds a GDP Minutes Receptor bg Effector 2 a GTP Effector 1
Signal Termination, Downregulation and Reset to Basal State G-Receptor Kinase (GRK) RGS Seconds a GDP Minutes Receptor bg a GTP Effector 1
Signal Termination and Reset to Basal State Inactive Basal State Receptor Receptor P P P a GDP bg Phosphatase G-Receptor Kinase (GRK) Dephosphorylation And rebinding of Ga and bg. (minutes) RGS Seconds a GDP Minutes Receptor P P P bg Arrestin Only activated receptors are phosphorylated and downregulated. This effect is slow (minutes) compared to activation (seconds). During this perturbation from steady state, PI3K activation occurs, driving directional motility. a GTP Effector 1
Integral Feedback Control Analogous to model in Yi, Huang, Simon&Doyle 2000 PNAS 97, 4649 If we assume that only activated receptors are phosphorylated (and thus inactivated) and that the phosphatase that dephosphorylates the GPCR operates at saturation and is less active than the G-protein Receptor Kinase (GRK), then the model is analogous to integral control of bacterial chemotaxis receptors. Inhibition of active chemotaxis receptors by demethylation is analogous to inactivation of active GPCRs by phosphorylation. This is a consequence of the fact that GRKs only phosphorylate receptors associated with active bg proteins. The rate of receptor phosphorylation is: dRP/dt = VPmax - VKmax(A)/(KK+A) (where A is the concentration of activated receptors, KK is the KM of the GRK for activated receptors, VPmax is the maximal activity of the phosphatase and VKmax is the maximal activity of the kinase, GRK ). Thus, the activity at steady state will be: Ast=KKVPmax/(VKmax-VPmax) This is the set point (y0 in the model above). y is defined as the difference between the activity at time t (y1) and the activity at steady state (y0). Thus, at steady state, y = 0.
t 0 Increased ligand binding acutely increases u and elevates y1 to a value above y0, giving a transient positive value for y (resulting in PI3K activation).At steady state, (y = 0) the rate of phosphorylation and dephosphorylation are equal. If one assumes that GRK only acts on active receptors (whether or not ligand is bound) then the netrate of phosphorylation at any instantaneous time will be directly proportional to y (the transient excess in active receptors over the steady state value). When y = 0 phosphorylation and dephosphorylaiton cancel out. The fraction of phosphorylated receptors (x) at any time t is then determined by the number of receptors in the phosphorylated state at time zero, x0 (e.g. prior to the perturbation due to increased ligand binding) plus the number of receptors that get phosphorylated during the interval in which the system was perturbed. This latter term is the integral from the time at which the perturbation (e.g. ligand unbinding) occurred t=0 to time t of ydt. So x(t) = x0 + ydt Notice that y can be + or - depending on whether ligand decreases or increases. Thus dx/dt = y = k(u-x) - y0 At steady state, dx/dt=y=0 and y1=y0 Notice that since k and y0 are constants, an increase in u (rapid binding of ligand) is ultimately offset by a slow decrease in x so that at steady state k(u-x) = y0.
Regulation of protein-Tyr kinases Tyr-P Kinase Kinase Kinase P-Tyr Tyr-P P-Tyr PI3K SH2 Autopho-transphorylation of low activity monomeric protein kinases in the ligand-induced dimer stabilizes the active state of each monomer, allowing further transphosphorylation at sites that recruit signaling proteins.
Tyr-P Kinase Kinase P-Tyr SHP2 Tyr-P P-Tyr SH2 SH2 containing phosphoTyr phosphatases (e.g. SHP2) are preferentially recruited to activated receptors and play a dual role of transmitting additional signals (Ras activation) and turning off receptors.
Prior to stimulation, protein-Tyr kinases have floppy activation loops (region containing Tyr 1157, 1162 and 1163 of the insulin receptor). As a consequence the enzyme has a low probability of being in the active conformation (~1%). Despite this low activity, when brought in proximity with a another low activity Tyr kinase (due to growth factor binding), cross-phosphorylation of respective activation loops can occur. Phosphorylation of the residues on this loop stabilizes the active conformation of the protein giving a ~100 fold increase in activity.
Activated Insulin Receptor Peptide substrate
Integral Control of Receptor Protein-Tyr Kinases The preferential dephosphorylation of activated Protein-Tyr kinases by SH2-containing phosphatases provides a potential mechanism for integral control. In response to an acute elevation in the level of ligand, the receptor will be rapidly activated, but in the continuous presence of the ligand, the phosphatase will ultimately return the kinase to a steady state activity that is determined by the affinity of the phosphatase for the activated kinase, the Vmax of the phosphatase and the Vmax of the kinase for transphosphorylation. Analogous to the set point for bacterial chemotaxis receptors one can show that: Ast = KM-SHP2VKinmax/(VSHP2max - VKinmax) This simplified system does not reset to the same steady state as prior to receptor stimulation since VKinmax is dependent on receptor ligation. Modeling predicts an overshoot followed by return to a steady state that depends on ligand occupation. This is in agreement with observations at intermediate times (0 to 30 min.) following PDGF stimulation Exclusive ubiquitinylation, of activated protein-Tyr kinases (due to SH2-containing E3 ligases (e.g cbl)), leads to receptor internalization, providing a second mechanism of longer term shut-off that also models as integral feedback control.
Integral Control of PI3K PI3K, when activated, phosphorylates lipids at a high rate but also autophosphorylates (on regulatory and catalytic subunits) at a slow rate, leading to inactivation. Assuming that the phosphatase that dephosphorylates PI3K is saturated by substrate, this could also lead to integral control of this enzyme.
Parallels between low molecular weight G protein (Ras, Rac Rho) regulation and heterotrimeric G protein regulation GDP/GTP Exchange Factor (GEF) activate: analogous to GPCR GEF (SOS) Ras-GDP Ras-GTP Effector slow basal Effectors such as Raf (Ser/Thr kinase) or PI3K bind to activated Ras GAP GTPase Activating Protein Analogous to RGS Heterotrimeric and low molecular weight GTP binding proteins have been retained and expanded during evolution because they have unstable activated states and can spontaneously return to inactive states. Inactivation can also be accelerated by GAPs.
Signal Transduction in Eukaryotic cells is usually initiated by recruitment of signaling proteins to the plasma membrane. We have discussed three major mechanisms for acute and reversible protein relocation in response to cell stimulation. These mechanisms have the potential to amplify small signals. More importantly, recruiting signaling proteins from a 3-dimensional space (cytosol) to a 2-dimensional space (membrane) provides a mechanism for facilitating unfavorable multimeric interactions. PI-3,4,5-P3 GTP-Ras P-Tyr PH AKT SH2 Ras GEF Activation of GTP-binding proteins Protein phosphorylation to create docking site Generation of lipid second messengers Membrane Membrane Membrane PI-4,5-P2 GDP-Ras Tyr PI3K Stochiometric Small Amplification Large Amplification
Sarraste Plasma Membrane The PH domain of BTK binds the head group of PI-3,4,5-P3 and crystallizes as a dimer with the two binding pockets on the same surface. However, in solution, it behaves as a monomer.
Moving signaling proteins from the three dimensional environment of the cytosol to the two dimensional environment of the plasma membrane decreases the entropy difference between a monomeric and dimeric state. Many signaling proteins may have evolved very weak free energies of homo or hetero-dimerization to insure that dimerization only occurs when confined on a two dimensional surface. Signal Dimers Monomers Membrane
Pólya's Random Walk Constants http://mathworld.wolfram.com/PolyasRandomWalkConstants.html Let p(d) be the probability that a random walk on a d-D lattice returns to the origin. Pólya (1921) proved that (1) p(1) = 1; p(2) = 1 but (2) p(d) < 1 for d > 2. Watson (1939), McCrea and Whipple (1940), Domb (1954), and Glasser and Zucker (1977) showed that (3) p(3) = 1 - 1/u(3) = 0.340537…. where u(3) = 3/(2p)3 p p p -p -p -p _____dxdydz______ 3-cosx-cosy-cosz Finch, S. R. "Pólya's Random Walk Constant." §5.9 in Mathematical Constants. Cambridge, England: Cambridge University Press, pp. 322-331, 2003. Domb, C. "On Multiple Returns in the Random-Walk Problem." Proc. Cambridge Philos. Soc. 50, 586-591, 1954. Glasser, M. L. and Zucker, I. J. "Extended Watson Integrals for the Cubic Lattices." Proc. Nat. Acad. Sci. U.S.A. 74, 1800-1801, 1977. McCrea, W. H. and Whipple, F. J. W. "Random Paths in Two and Three Dimensions." Proc. Roy. Soc. Edinburgh 60, 281-298, 1940. Montroll, E. W. "Random Walks in Multidimensional Spaces, Especially on Periodic Lattices." J. SIAM 4, 241-260, 1956. Sloane, N. J. A. Sequences A086230, A086231, A086232, A086233, A086234, A086235, and A086236 in "The On-Line Encyclopedia of Integer Sequences." http://www.research.att.com/~njas/sequences/. Watson, G. N. "Three Triple Integrals." Quart. J. Math., Oxford Ser. 2 10, 266-276, 1939. Eric W. Weisstein. "Pólya's Random Walk Constants." From MathWorld--A Wolfram Web Resource. http://mathworld.wolfram.com/PolyasRandomWalkConstants.html
Further websites for random walks http://www.rwc.uc.edu/koehler/biophys.2ed/java/walker.html http://www.krellinst.org/UCES/archive/modules/monte/node4.html