Download
1 / 39
390 likes | 531 Views
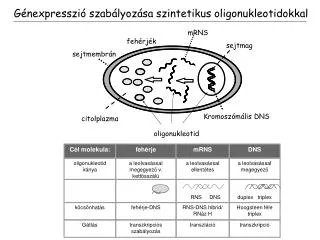
Cél molekula:. fehérje. mRNS. DNS. oligonukleotid iránya. a leolvasással megegyezõ v. kettõsszálú. a leolvasással ellentétes. a leolvasással megegyezõ. mRNS. RNS DNS. duplex triplex. fehérjék. sejtmag. sejtmembrán. köcsönhatás. fehérje-DNS. RNS-DNS hibrid/ RNáz H.

E N D
Cél molekula: fehérje mRNS DNS oligonukleotid iránya a leolvasással megegyezõ v. kettõsszálú a leolvasással ellentétes a leolvasással megegyezõ mRNS RNS DNS duplex triplex fehérjék sejtmag sejtmembrán köcsönhatás fehérje-DNS RNS-DNS hibrid/ RNáz H Hoogsteen féle triplex Gátlás transzkripciós szabályozás transzláció transzkripció Kromoszómális DNS citolplazma oligonukleotid Génexpresszió szabályozása szintetikus oligonukleotidokkal
Triplex képződésének kölcsönhatásai homopurin homopirimidin szekvenciáknál
Antiszensz oligonukleotidok lehetséges célszekvenciái a mRNS-en • exon-intron határhoz, slicing gátlás • Az 5’ sapka régióhoz • Transzlációs iniciációs régióhoz • Génen belüli régióhoz • Kapcsolódó oligonukleotidok Elsődleges hatásmechanizmus: Az endogén RNázH leemészti a DNS:RNS hibrid RNS szálát
Oligonukleotid szintézis:foszforamidit módszer B B 1 1 Ac O O O 2 O O H H O O O O hordozó hordozó B 1 B 1 O O O O di-MeO Tr A c O O 2% DCA/DCM O O di-MeO Tr hordozó hordozó < 1 % B 2 O O tetrazol di-MeO Tr O N újabb ciklus O P N N C O > 99 % B 2 B 2 O O di-MeO Tr O O O I / H O B 2 2 1 di-MeO Tr O O B O 1 O O O P O O N C O O P O O N C O O hordozó hordozó
Oligonukleotid szintézis:foszforamidit módszer B B 1 1 Ac O O O 2 O O H H O O O O hordozó hordozó B 1 B 1 O O O O di-MeO Tr A c O O 2% DCA/DCM O O di-MeO Tr hordozó hordozó < 1 % B 2 O O tetrazol di-MeO Tr O N újabb ciklus O P N N C O > 99 % B 2 B 2 O O di-MeO Tr O O O I / CS2 B 2 1 di-MeO Tr O O B O 1 O O O P O O N C S O P O O N C O O hordozó hordozó
Oligonukleotid szintézis: H-FOSZFONÁT MÓDSZER H lehet Me is Előny: az oxidációt elég a végén elvégezni,vagy – igény szerint – el sem kell töltés nélküli molekula Könnyebben átmegy a sejtfalon/membránon
PEPTID NUKLEINSAVAK (PNA) Nincs cukorfoszfát gerinc, ehelyett peptid kötéssel összekapcsolt lánc van Specifikus Stabil PNA T10-Lys TTTTTTTTTT-Lys PNA SV40-Lys ATTTTCTTCATTTTTTCTTC-Lys Ma már külön könyv van a PNA- alkalmazásairól
MUTAGENEZIS - in vivo gének elrontására,vagy módosítására • Random mutagenezis • találomra létrehozott mutációk • mutagén anyagok: UV, kemikáliák, radioaktív sugárzás • PCR: a hőstabil polimerázok átírási hűsége rosszabb • mutagén törzsek • transzpozon mutagenezis Irányított mutagenezis Adott helyen - deléciók inszerciók léterhozása - pont mutációk létrehozása
Gének irányított szétroncsolása interpozon mutagenezis Ar1 oriV oriT oriV: szűk gazdaspecificitás Ar2 Ar: antibiotikum rezisztencia Ar2 Ar1 oriT ori Ar2 vad típus mutáns poláris hatás
Deléciós mutagenezis a leolvasási keret sértése nélül Ar1 oriV oriT oriV: szűk gazdaspecificitás Ar: antibiotikum rezisztencia Ar1 ori oriT vad típus mutáns poláris hatás ?
Gének irányított mutagenezise a dut/ung rendszer alapján pontmutációk! Dut: dUTPase Ung: uracil-N-glikoziláz Ne épüljön be dezoxiuracil A DNS-be in vitro szintézis, Klenow
FEHÉRJE TERMELTETÉS VAN GÉNÜNK: Legyen az prokarióta vagy eukarióta természetes vagy szintetikus vad típusú vagy mutáns
FEHÉRJE TERMELTETŐ RENDSZEREK PROKARIÓTÁK E.coli ismert, Bármilyen fehérje, feltéve, ha - nem túl nagy - nem túl kicsi - nem túl hidrofób - nincs túl sok cisztein benne szekréciós rendszere minimális Bacillus subtilis jól ismert, ha nem is annyira, mint az E.coli van szekréciós rendszere EUKARIÓTÁK • emlős sejtvonalak • minden fajta fehérje termeltethető bennük • tranziens expressziós rendszerek viszonylag gyors, de korlátozott lehetôségek, • stabil expressziós rendszerek • hosszú, fáradságos optimalizálást igényel • élesztő • gyorsan szaporodó eukarióta sejtvonal, • sok ismeretanyag • viszonylag könnyű kezelhetőség • poszttranszlációs modifikációk lehetősége
Escherichia coli • ELŐNYÖK • óriási mennyiségű ismeretanyag • könnyű kezelhetőség, gyors növekedési sebesség, viszonylag olcsó médium • nagy mennyiségű biomassza gazdaságos előállítása • ismert szelekciós rendszerek • legtöbb expressziós rendszer • HÁTRÁNYOK • általában nem szekretál • a sejten belül redukáló atmoszféra diszulfid hidakkialakulása gátolt • az eukariota poszttranszlációs módosítások általában nem megvalósíthatóak • nem alakul ki a megfelelő aktiv szerkezet, rossz folding
STRATÉGIÁK TÚLTERMELTETÉSRE KONSTITUTÍV PROMÓTER INDUKÁLT TERMELTETÉS • a fehérje a teljes növekedési fázis alatt expresszálódik • nagy sejttömeg elérése után a fehérje visszanyerése • nem igazán használatos, toxicitási problémák miatt • a promoter represszált állapotban van a növekedés egy bizonyos fázisáig • valamilyen indukcióval derepresszió • további növesztés • fehérje visszanyerése • legáltalánosabban használt stratégia • toxicitás sok esetben megoldható
TSZE PROKARIÓTA EXPRESSZIÓS VEKTOR ELEMEI SZF GS Pr -35 -10 SD kódoló szekvencia TT STOP kodon UAAU UGA UAG -10 -35 TTGACAN17TATAAT START kodon AUG GUG UUG 5’ UAAGGAGGN(3-11) AR ORI Pr: promóter TT: transzkripciós terminációs szignál, SZF: szabályozó fehérje,, TSZE: transzlációt szabályozó elemek, SD: Shine-Dalgarno szekvencia, AR: antibiotikum rezisztencia, ORI: replikációs origo GS: gazdasejt
transzkripció transzláció A génexpresszió szabályozása DNS RNS FEHÉRJE transzripciós szinten poszt-transzkcripciós szinten (mRNS degradáció, protein stabilitás,stb.)
Különböző faktorok relatív hatása a fehérje termeltetésre E. coli-ban • Promóter indukálhatóság kb. 1000 x • Gazdasejt kb. 1000 x • A mRNS 5’ struktúrája kb. 100 x • Transzláció kb. 100 x • növekedési sebesség kb. 50 x(mRNS stabilitás)
túltermeltetésnél: ne legyen nagyon magas a kópiaszám toxikus fehérjék alapexpressziójának visszaszorítása “Runaway” plazmidok: indukálható kópiaszámú plazmidok
TRANSZKRIPCIÓ • Iniciáció • fehérje termeltetés során a legfontosabb • Elongáció mivel a gén belsejében van, nem manipulálható • Termináció a transzkripció befejezése, a transzkripciós apparátus túlterhelésének elkerülésére
NTD /` CTD CTD ? 5’ 3’ 3’ 5’ 5’ 3’ A transzkripció iniciációja prokariótákban sigma faktor holoenzim RNS polimeráz RNS polimeráz alegységek: , `, , , promóter 5’ 3’ 3’ 5’ k1 k-1 Ha k1 >> k-1, akkor a polimeráz holoenzim nagyon erősen kötődik a promóter szekvenciához, a polimeráz nehezen tudja elindítani a polimerizációs reakciót. Tehát a transzkript mennyiségecsökken, habár in vitro a promóter és a holoenzim közötti kapcsolat erős. k2 k-2 3’ 5’ 5’
A tökéletes70-függő promóter UP elem -35 Ext. -10 D +1 AAATAAAATTTTTAAn..nTTGACAnnn…nnnTGnTATAATnnattA 14nt 4-6nt 4-6nt 17nt As70ismeri fel Az aalegység kötődik
Különböző gének expresszióját különböző szigma faktorok befolyásolják tápelvonás, Stressz, Vasra éheztetés, sejtsűrűség, Felületi adházió Transzkripciós faktorok (Szigma) gén expresszió változás Morfológiai & fiziológiásváltozások Adaptáció / Védekezés
Enzim aktivitás mérés Western blotfehérje detektálása fehérje érése Riportergén riporter fehérje aktivitás promóter aktivitás mRNS detektálása transzláció transzkripció A vizsgált operon promóter A génexpresszió tanulmányozása különböző módszerekkel különböző szinteken
ELECTRO MOBILITY SHIFT ASSAY (EMSA) I. • más néven gél retardációs assay • a DNS-t, amely feltételezhetően fehérjét köt, megjelöljük • jelölés lehet végen, szálon belül, egyszálon, kétszálon, kötési kísérletben a DNS kétszálú! • a DNS ne legyen től hosszú 20-200 bp, minél hosszabb a DNS, annál több nem specifikus reackió történik • lehet tiszta fehérje vagy fehérje populáció Natív poliakrilamid gél-: fehérje nélkül, +: fehérjével összekeverve DNS - + - + jelölés DNS kötő fehérje nem kötődik kötődik összekeverés - + nem kötődik kötődik nem specifikusan kötődik
ELECTRO MOBILITY SHIFT ASSAY (EMSA) II. • a mobilitás eltolódás inkább a komplex méretét jellemzi, a kölcsönhatás errősségét nem • a kölcsönhatás erősségét a nem kötöt DNS, és a kötésben levő DNS jelének intenzitása jellemziminél kevesebb fehérje okoz erős jelet az eltolódott sávban annál erősebb a kölcsönhatás SUPERSHIFT ASSAY titrálás fehérje mennyiségre • ha van tippünk, hogy milyen fehérje kötődik • és van rá specifikus ellenanyagunk hozzáadott fehérje mennyiség nő - + nem kötődik kötődik ellenanyag is kötődik - + ++ a DNS - fehérje kölcsönhatás erősségéről ad felvilágosítást, nem a képződött mRNS mennyiségéről -: fehérje nélkül, +: csak fehérje hozzáadása, ++: fehérje és ellenanyag hozzáadása
* * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * DNázI DNázI FOOT PRINT I. DNS hossz 100 – 200 bp DNS jelölés szigorúan a végén, szálspecifikusan mind a két jelölt szállal meg kell csinálni a kötési kísérletben a DNS kétszálú denaturáló poliakrilamid gélelektroforézis Maxam-Gilbert létra DNázI hasítás G A+G C+T C D *
* * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * DNázI FOOT PRINT II. DNázI hasítás-: fehérje nélkül+: fehérje jelenlétében Maxam-Gilbert létra G A+G C+T C D- D+ hiperszenzitív hely (HSH) DNázI HSH
* * * * * * * * Me Me Me * * * * * * * * METILÁCIÓS INTERFERENCIA DNS jelölés szigorúan a végén, szálspecifikusan a DNS-t Maxam-Gilbert szerint dimetilszulfáttal (DMS) metiláljuk mind a két jelölt szállal meg kell csinálni a kötési kísérletben a DNS kétszálú denaturáló poliakrilamid gélelektroforézis piperidinhasítás MeG-nél Maxam-Gilbert létra G A+G C+T C G G G G G piperidin DMS G G G Me G csak a jelölt szálon szemléltetve
* * * * * * * * * * * METILÁCIÓS INTERFERENCIA II. piperidinhasítás MeG-nél Maxam-Gilbert létra modell: PM és NKmintázat K mintázat G A+G C+T C PM K NK tegyük fel, hogy ezek a G-k esszenciálisak a kötéshez DMS EMSA - + K: kötött NK: nem kötött kivágás, izolálás piperidin hasítás denaturáló PAGE PM: protein mentes
MS? UV KERESZTKÖTÉS (CROSS LINKING) Ismeretlen DNS kötő fehérjék molekulatömegének becslésére 1. A DNS szálat szálon belül jelöljük, és Br-dUMP-t építünk be (a Br-dUMP nem zavarja a kötődést) minta marker : Br SDS-PAGE jelölt fehérje DNázI kezelés EMSA UV fény kivágás, izolálás a fehérje DNS komplex kötés kovalens lesz
A mRNS mennyiségének meghatározása • Northern-blot és hibridizáció • reverz-transzkripció kapcsolt kvantitatív PCR • korrekt, de belső összehasonlító standardokat igényel
transzkripcionálisan aktív sejt vagy sejtmagi extraktum + ATP, CTP, 32P-UTP, In vitro transzkripció RegulátorG mentes kazetta régió (3-400 bp) Promóter a képződött radioaktív RNS analizise denaturáló PAGE-n. csak a fenti 3-400 bp-os fragmentum fog látszani, a többi tartalmaz G-t ezek transzkirpciója leáll mennyiségi becslés: denzitometriásan, vagy PhosphorImager-rel elsősorban transz szabályozó elemek hatásának vizsgálatára alkalmas
Riporter gének alkalmazása Regulátor régió riporter gén (pl. lacZ) Promóter SEJT -galaktozidáz (LacZ) -galaktozidáz aktivitás mérés feltételezi, hogy nincs poszt-transzkripciós szabályozás, és az enzim specifikus aktivitása is minden mintára egyforma aktivitás mérés kiküszöbölése GFP: Green Fluorescent Protein, gerjesztve floureszkál a kapott aktivitást illetve szignált normálni kell a sejt vagy protein mennyiségére