Download

1 / 31
330 likes | 723 Views
Organogenesis of secondary lymphoid tissues (SLT). Cytokines, chemokines and cell adhesion molecules. Lymphoid tissue: primary and secondary sites. Primary lymphoid tissue. Bone marrow, and foetal liver. Thymus (absent in nude mice – whn transcription factor mutation).

E N D
Organogenesis of secondary lymphoid tissues (SLT). Cytokines, chemokines and cell adhesion molecules.
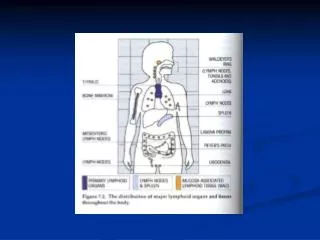
Lymphoid tissue: primary and secondary sites. Primary lymphoid tissue. • Bone marrow, and foetal liver. • Thymus (absent in nude mice – whn transcription factor mutation). • Secondary lymphoid tissue (SLT). • Spleen • - Developmentally separate from other SLT. Distinct genes involved: • hox11, Bapx1, Wilms tumour suppressor (WT1), capsulin • - Architecture often disrupted when LN and PP lost by mutation but • the spleen is still there. • Lymph nodes (LN). • - Mucosa-associated lymphoid tissues (MALT). • - Bronchial associated lymphoid tissue (BALT). • - Nasopharyngeal-associated lymphoid tissue (NALT). • - Gut associated lymphoid tissue (GALT). • Peyer’s patches (PP). • Lymphoid clusters.
Function of secondary lymphoid tissue. • To permit efficient interactions between antigen, antigen-presenting cells, lymphocytes and other regulatory cells. • To provide a controlled environment for the development of immune responses.
Lymph node location. The lymphatic vasculature drains tissue fluid, cells and antigens from most tissues, through LNs and back into blood via thoracic duct. Axillary – armpit. Brachial – On bicep, underneath pectorals. Deep cervical – Behind salivary gland. Superficial cervical – In front of salivary gland. Inguinal – Adherent to skin of groin. Lumbar – Behind split of abdominal aorta. Mediastinal – Thymic region. Mesenteric – Mesentery of small intestine and pancreas. Popliteal – Behind the knee. Pancreatic – Between pancreas and stomach. Renal – Between aorta and kidneys. Sacral – In front of the split of the abdominal aorta. Sciatic – Below sciatic nerve. Facial – draining the face. They are always in the same place !
HEV Blood High endothelial venules: HEV Afferent lymphatic: antigen, APCs, T cells and tissue fluid. Network of channels connect lymphatic to blood vessels. Pathways for antigens, other peripheral molecules, and cells. Paracortical cords act as T cell channels. Cortex Paracortex Dendritic cells. Medulla Efferent lymphatic: cells, antibodies and tissue fluid. Neonate lymph node structure. B B T T T T Plasma cells Macrophages
Antigen delivery different from LNs. Use M cells. M cells Follicle-associated epithelium Efferent lymphatic vessel To mesenteric LN. Neonate Peyer’s Patch Structure. Gut lumen Follicle Villus HEV B B T T Dendritic cells. NALT has a related structure.
DCs in T and B cell areas. Red Pulp Marginal sinus Antigen and cell entry B T Conduits lined with fibroblasts Central arteriole B T PALS Marginal zone (MZ) T White Pulp B Adherent MZ B cells Macrophages Spleen
Time line of the development of lymphoid organs. Different parts of the secondary lymphoid tissue system develop at different times. Environment induces further enlargement and development after birth. First PP forms at border of duodenum and ileum, and they are then generated successively, one by one towards the lower intestine at regular intervals, although the final number is variable.
LT LT-R-Ig LT-R Make a LN or PP Revealed by in situ analysis (limited by embryonic LN size) or by inhibiting or activating various receptors at different points of embryogenesis (i) blocking lymphotoxin(LT) with a LT-R-Ig fusion protein. Stops PP and LN development. (ii) using an LT-R agonistic antibody in LT null mice. LNs and PPs are rescued. (iii) blocking IL7Ra with an antibody, blocks PP formation. But if PP has started, it finishes. Stops LN/PP development .
IL7? LT gene deleted IL7Ra Mimics LT Inhibits IL7Ra Make a LN or PP LNs and PPs are rescued Revealed by in situ analysis (limited by LN size) or by inhibiting or activating various receptors at different points of embryogenesis (i) blocking lymphotoxin(LT) with a LT-R-Ig fusion protein. Stops PP and LN development. (ii) using an LT-R agonistic antibody in LT null mice. LNs and PPs are rescued. (iii) blocking IL7Ra with an antibody, blocks PP formation. But if PP has started, it finishes. LT-R No LNs or PPs Prevents PP formation The time at which these inhibitors or activators are added determines which LNs and PPs are affected.
Mutant mice with defective lymphoid organogenesis – Essential secondary lymphoid tissue (SLT) genes. Gene deletion (‘knock-outs’) has revealed major molecular players. Cytokines and receptors. LTa, LTb, LTbR, TNFR, TranceR, Trance, IL7, IL7R Common cytokine receptor g-chain (gc). Signal transduction molecules and transcription factors. Jak3, Nik, IKKa, rel-a, rel-b, traf6, Id2, Ror-g, Ikaros, NFkb2. Chemokines and receptors. CCR7, CXCR5, CXCL13. Other essential genes may be missed if they give a lethal embryonic phenotype (CXCR4).
HOW DO THESE MOLECULES CO-ORDINATE LYMPHOID TISSUE DEVELOPMENT? Peyer’s patches as a model.
Anti-VCAM-1 stain Adachi Int Immunol. 9, 507. Stromal organiser cell (VCAM-1+/ICAM-1+) IL7Ra+ cell Lymphatic Mesenchyme Blood vessel Step 1: Formation of the early Peyer’s Patch ‘organiser’. Clustering of stromal ‘organiser’ cells around a lymphatic vessel. - visualised with antibodies to VCAM-1 or ICAM-1 (Cell Adhesion Molecules). May be directed by IL7Ra-expressing cells. Distribution? Anti-mesenteric side. How do they know where to start?
chemokines Stromal organiser cell (VCAM-1/ICAM-1) IL-7Ra+ inducer cell ? MAdCAM-1+ HEV LTICs (inducers) Lymphatic Mesenchyme Blood vessel Step 2: Colonising the developing SLT with‘inducers’. A subset of blood cells migrate out of the blood vessel, between the cells of the HEV, and into the developing SLT. These are specialised cells termed ‘Lymphoid Tissue Inducing Cells’ (LTICs) or ‘inducers’. May be same as early IL7Ra+ cells. Further clustering with stromal cells. Accompanied by new blood vessel supply – angiogenesis. ECs express markers that allow for blood cell homing.
INDUCER Stem cell Multipotent Precursor. Restricted blood borne SLT ‘inducer’. Are some of the essential SLT genes involved in making the ‘inducer’ cells? LN and PP recovered by injecting normal LTIC inducer cells into mice lacking genes involved in generating the LTICs.
TRANCE RANKL OPG On inducer cells. On organiser cells. TRANCER RANK (on inducer cells) What do the ‘inducers’ bring with them? Members of the TNF superfamily – central role of Lymphotoxin. So, if a receptor is required for LN and PP formation, maybe molecules involved in signalling from that receptor are also required………
Signalling through LTbR on ‘organiser’ cells. IKKa IKKb IKKg Cytoplasmic retention proteins.
Removing the LTbR signal transduction pathway reduces chemokine expression by organiser cells, so fewer LTIC inducers are attracted. * * *
Rolling Firm Adhesion INTEGRINS Extravasation SELECTINS CHEMOKINES Getting the inducer cells to the right place. Chemokines direct the homing and migration of LTICs during development, and control leukocytes in the adult. Blood flow - Three different chemokines Tissue .
CXCL13/BLC CXCL12/ SDF1 CD4 IL7Ra CXCR5 CXCR4 a4b1 integrin (VCAM-1 receptor) CCL19/ELC a4b7 integrin (MAdCAM-1 receptor) CD11c CCR7 L-selectin (PNAd receptor) CCL21/SLC Chemokine receptors and integrins on inducer cells. INDUCERCELL Chemokine production by organiser cells causes…… Activation of integrins to adhere inducers to endothelial cells at these sites, via MadCAM-1 (in PP and early LN), or other integrin ligands (VCAM-1). Null mice: Cooperation between CXCR5 and CCR7 for LN and PP development. Chemokines are not just chemoattractants………
Honda et al (2001) J. Exp. Med., 193, 621. CXCR5/CCR7 co-operation in LN/PP development. IL7R-/- Most LN present. MLN present but smaller. No PP. CXCR5-/- or CXCL13-/- Some LN present. MLN present but badly organised. Few PP CCR7-/- Most LN and PP present, but badly organised. CXCR5-/-CCR7-/- No PPs or LNs except MLN, which is badly organised. CXCL13-/-IL7Ra-/- No PPs or LNs, including MLN. Some LN/PP use CXCL13 only. Some use CXCL13 andCCL19/21. MLN use CXCL13 and IL7Ra ligand. MLN – Mesenteric LN.
LTa1b2 Stimulation through CXCR5 and CCR7: not just migration. CXCL13 through CXCR5, CCL19/21 through CCR7 causes the LTICs to: - up-regulate Lymphotoxin LTa1b2expression. - adhere to VCAM-1 by activating a4b1 integrin. Outcome: - Adhesion of inducer cells to stromal organiser cells. - Stimulation of LTbR. AND LTbR stimulation causes the up-regulation of the CXCL13 and CCL19/21 chemokines, and IL7. POSITIVE FEEDBACK LOOP……….
Model for early SLT organogenesis. LN Inducer IL7R ligands PP Organiser Cytokines, chemokines, and cell adhesion molecules.
EPITHELIAL CHANGES M cells Follicle Follicle-associated epithelium B HEV B B T T T Efferent lymphatic vessel COMPARTMENTALISATION What next in Peyer’s Patch development, once cell clusters form?
B cells – green T cells – blue Normal CCR7-/- CXCR5-/- CCR7-/-CXCR5-/- Lymphotoxin from B cells causes CXCL13 release from splenic stromal cells in developing follicle, attracting more B cells. Compartmentalisation probably driven by CXCL13, and CCR7 ligands, controlled by TNF family members – look in the spleen. Organisation of splenic B and T cell zones controlled by chemokines and TNF family members, even though the development of the spleen framework is normal. CXCR5 – B cellls. CCR7 – T cells. CXCR4 – T and B cells.
Induction of changes in the local epithelium. Electron micrographs of FAE (left) and M cell (right). Signals from the PP (from B cells?), induces formation of the Follicle Associated Epithelium (FAE) involved in antigen/pathogen uptake. M cells form and invaginations fill with lymphocytes (memory B and CD4+ T cells). FAE can also regulate cell influx into the PP region. - FAE makes chemokines: MIP-3a, which signals through CCR6, and CCL9, which signals through CCR1. - Mice without CCR6 lack DCs under FAE, and can’t mount good gut immune response. - Abs against chemokine CCL9 reported to reduce DC number.
Ectopic lymphoid tissue formation – tertiary lymphoid tissue. Inappropriate formation of lymphoid tissue can occur in many chronicinflammatory diseases. • Autoimmune diabetes. • Rheumatoid arthritis. • H. pylori (stomach) B. burgdorferi (skin) infection. • Hashimoto’s thyroiditis. • Sjogren’s syndrome. Important for strong responses to autoantigens and loss of self-tolerance. Therapeutic potential in disrupting these structures? Transgenic expression of CXCL13/BLC or CCL21/SLC or LTa/b in the pancreas, induces the formation of lymphoid tissue in this tissue. • The chemokine-induced formation of lymphoid tissue in these experiments is dependent on LTa1b2 and LTbR.
B cells T cells DCs HEVs SLC Accumulate injected labelled B cells Transgenic expression of BLC/CXCL13 in the pancreas using the Rat insulin promoter.
References. Some of the key papers: • Mebius (2003). Nat Rev Immunol., 3, 292. • Several excellent reviews in Immunological Reviews issue 195 (2003). • Ansel and Cyster (2001) Curr. Opin. Immunol. 13: 172. • Debard (1999) Sem. Immunol., 11:183. • Owen (1999) Sem. Immunol., 11:157. Pictures from: • Honda (2001) J. Exp. Med., 193: 621. • Adachi (1997) Int. Immunol., 9:507. • Hashi (2001) J. Immunol., 166: 3702. • Neutra (2001) Nat. Immunol., 2: 1004. • Finke (2002) Immunity, 17, 363. • Luther (2003) J Exp Med 197 1191. • Ohl (2003) J Exp Med 197, 1199.