Download
1 / 40
900 likes | 5.25k Views
Gastrulation is the first step of morphogenesis. Morphogenesis is the process whereby individual cells undergo complex movements that generate the organ rudiments. Gastrulation generates the three basic germ layers from which organs arise.

E N D
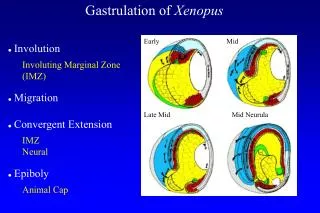
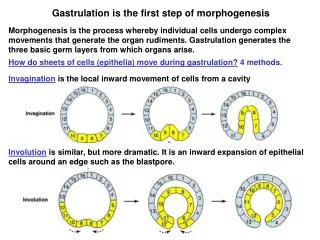
Gastrulation is the first step of morphogenesis Morphogenesis is the process whereby individual cells undergo complex movements that generate the organ rudiments. Gastrulation generates the three basic germ layers from which organs arise. How do sheets of cells (epithelia) move during gastrulation? 4 methods. Invagination is the local inward movement of cells from a cavity Involution is similar, but more dramatic. It is an inward expansion of epithelial cells around an edge such as the blastpore.
Convergent extension is elongation of an epithelium in one direction while it shortens in the other direction (stretching taffy). The cells can keep their relative positions and elongate or they can interdigitate. Epiboly is spreading movement of an epithelium to a deeper or thinner layer.
How do individual cells move during gastrulation? 4 basic types of cell movement lead to the changes in epithelial sheets that characterize gastrulation. 1. Migrationis the movement of an individual cell over other cells or a substrate. 2. Intercalationis wedging of cells between their neighbors. Lateral intercalation involves lateral movements of cells in the same layer between one another = convergent extension. Radial intercalation involves wedging of 2 different layers. This process often leads to epiboly, the surface area of the epithelium increases while the thickness decreases.
3. Ingression is the movement of individual cells from an epithelium into an embryonic cavity. 4. Shape changes are coordinated changes in cell shape that cause an epithelium to invaginate, buckle or undergo convergent extension.
Embryonic cells are broadly classed as epithelial or mesechymal Epithelial cells are well-differentiated. They compose skin and line the body cavities (ie, the digestive tract). They are polarized. Their apical surface faces out and their basal surface rests on the basement membrane (extracellular matrix that supports cells). Epithelial cells are closely connected with adjacent cells by specialized attachments including tight junctions, gap junctions, and desmosomes. Mesenchymal cells are poorly differentiated and have the potential to develop into many different tissues, including epithelial cells. They have a leading edge with lamellipodia, and a trailing edge. They are not connected to adjacent cells but they are in contact with the extracellular matrix.
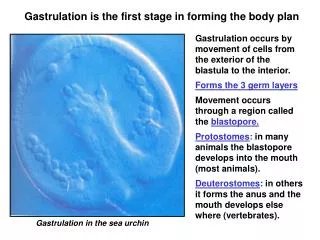
Gastrulation in sea urchins Primary mesenchymal cells: these cells change adhesive properties and the large micromeres start to migrate into the blastocoel as free mesenchymal cells (ingression). Mesenchymal cells are loose cells that can differentiate into many different organs. Archenteron: the primitive gut. The archenteron is formed in several stages. 1. the vegetal plate invaginates into the blastocoel, 2. It elongates by convergent extension. 3. It hooks up with the front and is pulled forward, and 4.Involution occurs with movement of cells around the blastopore and into the archenteron. What forces drive the process of gastrulation?
The primitive groove and pit are the site of gastrulation in birds Crossection of blastoderm (blastula in birds). Epiblast is the upper layer of epithelial cells, blastocoel is the space below the epiblast, and hypoblast is the lower layer of epithelial cells. Epiblast cells roll over the primitive ridge and involute into the groove. The cells lose contact with one another and migrate inwards by ingression = Mesoderm. 3 germ layers are established
Gastrulation in a 16 day old human embryo • The primitive streak and Henson’s node form just as in birds. • The cell movements are similar • Cells roll over the primitive ridge and into the groove • They ingress individually and move out to form discs with the 3 germ layers • The 3 layers also move laterally to form the extra embryonic endoderm and • mesoderm even though there is no yolk to digest. This is surprising • because mammalian embryos could gastrulate easily by invagination as sea • urchins (they have a placenta and no yolk). • Instead, gastrulation appears to recapitulate a pattern established by bird- • like ancestors and reptiles.
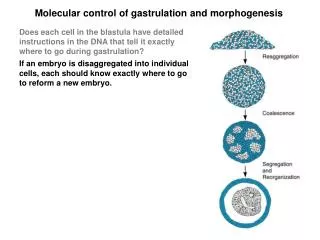
Molecular control of gastrulation and morphogenesis Does each cell in the blastula have detailed instructions in the DNA that tell it exactly where to go during gastrulation? If an embryo is disaggregated into individual cells, each should know exactly where to go to reform a new embryo. When this experiment is performed, a degree of reorganization occurs, but it is not complete. Embryoids: are slightly similar to embryos but they lack the real organization. Conclusions: 1. Genes impart only partial instructions for assembly of the embryo 2. Like cells all stick together, revealing distinct adhesive properties. 3. The relative positions of aggregates reflect the relative positions in the embryo (skin outside, heart inside).
Cell adhesion is the driving force in gastrulation When cells from an embryo are disaggregated and recombined, they can be readily ranked according to their ability to form the central portion. (chondrocytes > heart cells > liver cells is the hierarchical order) Differential adhesion hypothesis: the cell type with maximal adhesiveness (chondrocytes) will form a core that is surrounded by concentric spheres of cells with progressively lower adhesiveness. Cell adhesion can be measured by the ‘pancake test’. When aggregates of different cell types are subjected to a flattening force (centrifugation to induce a centrifugal force), the cells that adhere most tightly form a ball, while those that adhere more loosely form a flatter, pancake structure. Cell adhesion is a major factor that regulates aggregation of like cells and controls position during morphogenesis. What regulates how tightly or loosely cells attach?
Cells adhere by cell junctions, cell adhesion molecules, or substrate adhesion molecules Cell junctions:large, complex structures that form slowly but generate very strong and durable connections (tight junctions, desmosomes, and gap junctions). Cell adhesion molecules (CAMs):single molecules that traverse cell membranes and allow cells to adhere to one another. Adhesions form quickly, they are selective, but they are relatively weak in comparison to cell junctions. Substrate adhesion molecules (SAMs): a group that consists of extracellular matrix molecules and matched receptors that are expressed on the cell surface.
CAMs firmly anchor adjacent cells to the cytoskeleton Cell adhesion molecules (CAMs) are glycoproteins with 3 major domains: The extracellular domain allows one CAM to bind to another on an adjacent cell. The binding can be to the same type of cell (homotypic) or to a different cell type (heterotypic). The transmembrane domain links the CAM to the plasma membrane through hydrophobic forces. The cytoplasmic domain is directly connected to the cytoskeleton by linker proteins. This anchoring is important to prevent lateral diffusion of adhesion molecules in the membrane. Three major types of CAMs are immuno globulin-like CAMs, cadherins, and lectins.
Neural cell adhesion molecule is typical of immunoglobulin (IgG)-like CAMs N-CAM was one of the first to be discovered. The extracellular domain has IgG like repeats that are thought to allow binding to other N-CAMs by interdigitation between loops. Insects have IgG CAMs but no IgG. Thus, IgGs may have evolved from IgG-like CAMs. Polysialic acid region (PSA): 3 long carbohydrate chains with negative charge are attached to the 5th loop. The overall charge varies on different N-CAMs. Large PSA regions induce a large negative charge which repels cells (embryonic cells during gastrulation). Small PSA regions allow attachment due to low charges, and these are common on adult cells.
Cadherins mediate calcium-dependent cell adhesion Cadherins are the most prevalent CAMs in vertebrates. They are rapidly degraded by proteases in the absenceof Ca++. There are 4 major types: E cadherins in epithelial cells P cadherins in placenta N cadherins in neural tissue L cadherins in liver Each associates with its own type. 125 kD transmembrane glycoproteins that bind homotypically using the first 113 AA The differences in cadherin expression are responsible for the differential adhesiveness seen in disaggregated tissue. Cells that express more cadherin = tissues that form a ball in the center of cell aggregates.
Integrins mediate adhesion to ECM Integrins are a family of transmembrane glycoproteins that are composed of 2 chains, a and b. There are 40 different types of a chains and 8 types of b chains that can combine to form a large number of different integrin molecules. The a chain has binding sites for Ca++ and Mg++ which are needed for integrins to adhere. The 2 subunits form the site that binds to the RGD domain on ECM. The cytoplasmic tail of integrins is connected to a linker protein that connects to the cytoskeleton. A bridge from ECM to cytoskeleton.
CAM expression during gastrulation is correlated with cell fate Fate map: it is possible to predict which parts of the blastula will develop into specific structures after gastrulation. Expression map of CAMs: it is possible to localize expression of CAMs using in situ hybridization and immunostaining of the blastula. Cells with different fates express different CAMs. Cells destined to become neural tissue express high levels of N-CAM. Cells destined for epidermis express E-cadherin. The respective cell adhesion molecules are expressed before the cells actively start to form the adult tissue. This suggests that CAM expression is important in fate determination. During gastrulation, cells go where their CAMs lead them
Changes in cell adhesion are important for gastrulation Gastrulation in the sea urchin is initiated by specific changes in cell adhesion. One of the first steps is ingression of mesenchymal cells from the vegetal plate into the blastocoel to form the skeleton of spicules. The mesenchymal cells lose their adhesion to hyaline and the adjacent nonmesenchymal blastomeres. They start to increase adhesion to the basement membrane and material within the blastocoel. These changes can be measured by isolating specific cells and testing adhesion in culture. E-cadherin is lost from the ingressing cells due to endocytosis of specific areas where it was expressed. Levels of b-catenin are also reduced on these cells.
Fibronectin on the inner roof of the blastocoel is critical for gastrulation Immunostaining of the blastocoel shows that fibronectin was expressed in abundance on the inner roof. Fibronectin binds to integrins on the membrane. Neutralizing antibody to fibronectin was injected into the blastocoel to test the role of fibronectin. This aborted gastrulation. Since no epidermal cells could migrate into the blastopore, many cells accumulated on the surface, forming deep folds. If an unrelated antibody was injected, there was no inhibition. Fibronectin binds integrins through an RGD sequence. Similar results were obtained by injecting the tripeptide RGD. Furthermore, blocking the integrin receptor with injected antibodies also inhibited gastrulation. Fibronectin is important for contact guidance of migrating cells during gastrulation.
PC12 cells resemble chromaffin cells which can differentiate into neurons. When they convert to the neural phenotype they express N-CAM and N-cadherin on their cell surface. When PC12 cells are grown on cells that do not express N-CAM or N-cadherin (3T3 cells) they retain the undifferentiated chromaffin phenotype. If the PC12 cells are grown on the same cells that have been transfected with N-CAM or N-cadherin genes, they convert to the neuronal phenotype. They form long dendrites and express neuronal genes. Differentiation is accompanied by opening of calcium channels
Organogenesis in humans is essentially complete after 6-8 weeks The 5 week old human embryo has a head with rudiments of eyes, ears, and brain. It also has a trunk with tail and limb buds. The structures develop rapidly and the organs are basically formed before 8 weeks. The period of histogenesis, when cells acquire functional specialization, then begins and continues throughout development = fetus. Organogenesis involves many of the same cell behaviors that occur during gastrulation.
Neurulation is of scientific and medical interest Spina bifida: one of the most common birth defects in humans. Due to defects in closure of the neural tube and malformations of brain and spinal cord. There are several forms that differ in severity: Spina bifida occulta: the mildest form is caused by failure of a vertebrate to fuse dorsally. It causes no pain or neurological disorder. It is very common and as many as 10% have this minor defect. The only sign of its presence may be a dimple or a tuft of hair normal spina bifida occulta
What forces cause closure of the neural tube? • Closure of the neural tube is mediated by 3 • effects: • Apical constriction of neural plate cells • Rapid anteroposterior extension • Cell crawling • 1. Apical constriction: a band of • microfilaments in the apical region of neural • plate cells contracts and causes cells to • assume a wedge shape. This causes the • neural plate to bend and form the hinge • regions. • Apical constriction occurs in the midline • throughout the neural plate. It also occurs in • the lateral folds to form the mediolateral • hinge sites.
Spemann’s famous organizer experiment Spemann originally thought that the donor dorsal lip of the blastpore differentiated into neural tube and structures of the embryonic axis. To prove this, he grafted dorsal lip blastopore tissue from a nonpigmented donor newt onto another gastrula from a heavily pigmented species. When the embryos developed he found that the embryonic axis was actually composed of pigmented recipient tissue. Only a small strip of donor tissue was present in the middle of the neural plate.
Spemann’s experiments were confirmed in birds • 3 conclusions about the organizer: • The dorsal lip of the blastopore developed according to its own fate. It gastrulated normally in the new location and formed notochord. • The graft dorsalized the host’s ventral mesoderm (converted gut tissue to kidneys and somites). • The graft acted as a neural inducer. It caused host ectoderm to form a neural plate and close to become a neural tube.
Axis induction by disinhibition Early experiments by Spemann studied how the organizer worked. If the cells of the dorsal lip were killed or crushed, the activity was still present. Activity could be mimicked partially by changes in pH or ionic strength. Disinhibition: the normal pattern for ectodermal development is neurulation. Ventral ectoderm escapes from this path by producing an inhibitor of neurulation. The organizer works by inhibiting the inhibitor. Cells cultured from ventral ectoderm differentiate as neurons only if cultures are sparse. Bone morhogenetic protein (BMP-4) is released by ventral ectoderm cells. It stimulates formation of ventral structures and inhibits neurulation if injected into embryos. If BMP-4 is blocked, dorsal structures replace ventral structures (dominant negative mutants or innactivate the BMP-4 receptor).
Spemann’s organizer (dorsal lip) inactivates BMP-4 Chordin and noggin: the dorsal lip of the blastopore produces 2 proteins that antagonize the action of BMP-4. Chordin and noggin each bind to BMP-4 and prevent it from binding to its receptor. Chordin and noggin have strong dorsalizing effects. They cause excessive head development and block ventral differentiation if injected into the embryo. What turns on chordin and noggin? Goosecoid: may be the master regulator that is similar to Spemann’s organizer. Expression of goosecoid is limited to the dorsal lip of the blastopore. It is turned on at the right place at the right time. Goosecoid encodes a transcription factor that can activate chordin gene expression. Work is continuing to explore how this gene functions.
Histogenesis is the process by which cells and tissues acquire functional specialization embryo fetus Cleavage gastrulation organogenesis histogenesis (2 weeks) (1 week) (4 weeks) (7 months) Organogenesis: the formation of organ rudiments to establish the basic body plan. Histogenesis: differentiation of cells within the organs to form specialized tissues. Tissues are composed of cells and extracellular material that perform a specific function. Each specific tissue develops mainly from one germ layer.
The spinal cord develops a dorsoventral pattern All nervous functions depend on development of complex connections between neurons. These circuits start to develop in the embryo. Alar plate:as neurons and glial cells accumulate in the mantle, they form ridges on either side of the neural tube (dorsal or afferent columns). These will develop into afferent nerves that conduct signals to the brain. Basal plate:accumulation of cells in the ventral region produces basal or efferent columns. These will develop into efferent neurons that carry signals to muscles and organs (motor neurons). The gray matter in the mantle layer is composed of cell bodies of neurons and the white matter in the marginal layer is composed of myelinated axons. The floor and roof plates are composed of glial cells. What causes this pattern?
Sonic hedgehog (shh) induces the dorsoventral pattern Sonic hedgehog (shh) is a gene that is expressed in the notochord at first and later in the floor plate. Mice that lack shh fail to develop floor plates in the CNS. Shh is a secreted glycoprotein that induces a gradient that is high near the floor plate and progressively lower in dorsal regions. Shh initially induces neural plate cells to form floor plate. Other signals from the dorsal ectoderm direct the dorsal columns and the roof plate. Different levels of shh appear to specify different types of neuron differentiation.
Different concentrations of shh induce distinct types of neurons In the developing spinal cord, the floor plate produces shh and creates a concentration gradient. Motor neurons develop closest to the floor plate, type 2 interneurons are next, followed by type 1 interneurons. To see whether the shh gradient is really important, isolated cells from neural tubes were cultured in various concentrations of shh. Cells were stained with antibody specific for floor plate, motor neurons, or type 1 or 2 interneurons. Bone morphogenetic protein released by the dorsal ectoderm and roof plate has an analogous function in generating the dorsal columns
Neural crest cells form a variety of tissues The fate of neural crest cells has been mapped by a number of techniques (radioactive tracers, transplants from pigmented species to albinos). There are 2 patterns of migration in the trunk region: Dorsolateral path: enter skin and form melanocytes Ventral path: form afferent neurons of dorsal route ganglia, sympathetic and parasympathetic ganglia, and adrenal medulla Neural crest cells help to form addition structures in the head such as bones, connective tissue, eyes, ears, and teeth. They also help to form blood vessels and connective tissue in the trunk
How do neural crest cells differentiate into many tissues? Pluripotency hypothesis: each neural crest cell has the potential to form any or all structures. Inductive signals from adjacent tissue determines their fate. Selection hypothesis: the neural crest contains a mixed population of predetermined cells. Each cell has only one possible fate and it migrates according to this fate. The real truth may lie between these two extremes. Clonal analysis: when individual neural crest cells are placed in culture, it is clear that a single cell can give rise to others that differentiate into multiple cell types (pigment cells and neurons). Premigratory cells have a wider potential than do the cells that have already started to migrate. They may become partially differentiated as they migrate.
What are the molecular signals that control differentiation of neural crest cells? Extracellular matrix (ECM): neural crest cells constantly extend filopodia to feel the ECM. Pieces of filter were placed in an embryo at the dorsolateral or ventral pathways. After the filters absorbed ECM, they were removed to a culture dish and allowed to interact with neural crest cells. Dorsolateral ECM induced melanocytes and yellow pigment cells. Ventral ECM induced neurons. No matrix allowed the cells to remain undifferentiated.
The otic placode forms the inner ear In humans, the otic placode appears by the third week on both sides of the rhombencephalon. The otic vesicle is induced by the underlying neural tissue in the rhombencephalon. It invaginates to form the otic pit and then pinches off to form the otic vesicle. Ganglion cells develop from its medial surface. Labyrinth:the otic vesicle expands unequally and constricts in other areas to form a complex shape. The cochlea develops to sense sound and the semicircular canals form to serve as an organ for balance and body position.
Formation of the eye involves reciprocal interactions Lens placode: the ectoderm invaginates in response to signals from the optic cup underneath. It then pinches off as a lens vesicle. Cells elongate to fill the vesicle and start to synthesize crystallins. Optic cup: forms from the neural tube by invagination. The opening (choroid fissure) closes forming a round optic cup, an extension of the brain. Optic stalk: connection to the brain that is filled with neurons to form the optic nerve. Reciprocal interaction: the lens induces the formation of the optic cup and the cup regulates formation of the lens. When the lens from a species with large eyes is transplanted, it induces an extra large optic cup and it also does not grow as large as usual.
Nasal placodes form the olfactory epithelium In humans, the nasal placodes appear after the 4th week. They are induced by the underlying telecephalon. During the fifth week, nasal swellings appear around the placodes which now become the two nasal pits. The pits start out far apart, but the 2 maxillary swellings grow large and push the pits to the center. The medial parts of the nasal swellings fuse to form part of the upper lip. Olfactory epithelium: the original lining of the nasal pit comes to rest on the roof of the nasal cavity. It forms the epithelium that senses smell and connects to neurons in the telencephalon. The nasal cavity becomes continuous with the pharynx. What can go wrong?
How does skin develop and differentiate? Epidermis: the largest derivative of ectoderm forms the outer layer of the skin. It is an epithelium and cells are connected by desmosomes and tight junctions. The epidermis consists initially of two layers: periderm is a temporary outer layer and the germanative layer lies below. The germanative layer is composed of stem cells that divide actively to produce differentiated progeny. The basal layer develops from the germanative layer (it contains stem cells in the adult). Spinous layer forms as cells are squeezed out of the basal layer. They become large and differentiate. Granular layer starts making keratin granules Cornified or horny layer is composed of dead cells that are filled with keratin It takes cells about 7 days for each cell to journey through the skin
Hair development in humans The underlying mesenchyme in skin forms a dermis, a layer of connective tissue just below the epidermis. The dermis induces a variety of epidermal structures depending on the species (feathers, hair, scales). Hair bud: hair formation begins as a small bud that that penetrates the dermis. It is induced by a group of mesenchymal cells and the hair bud then envelopes these cells to form a hair papilla. Hair follicle is the entire organ Sebaceous glands are induced to form on the side. The hair shaft is formed when the inner cells of the follicle start to differentiate and produce keratin in the form of hair. The continued production of keratin by the cells at the base of the shaft causes the hair to grow longer.
Mammary gland development in humans mimicks ancestral patterns In normal development of humans, only one pair of segments in the ridge survives, and the remaining precursors degenerate. In some individuals, the other segments of the mammary ridge fail to degenerate, so that accessory nipples or breasts are formed. Primates evolved from small creatures that nursed multiple offspring. An extended mammary ridge would have given human ancestors a survival advantage. Two breasts are obviously more adaptive for humans who normally have only one offspring at a time. Atavism: the occasional and abnormal persistance of a primitive adult feature in an evolved species (multiple mammary glands). It is easier to modify an older pattern of development than to develop a totally new pattern. This idea is a pervasive in developmental biology.