Download

1 / 28
300 likes | 4.39k Views
BIOL2007 - EVOLUTION AT MORE THAN ONE GENE SO FAR Evolution at a single locus No interactions between genes One gene - one trait REAL evolution: 10,000 - 100,000 genes producing mRNA linkage, a physical interaction mechanistic interactions in gene action. GENE INTERACTIONS

E N D
BIOL2007 - EVOLUTION AT MORE THAN ONE GENE • SO FAR • Evolution at a single locus • No interactions between genes • One gene - one trait • REAL evolution: • 10,000 - 100,000 genes producing mRNA • linkage, a physical interaction • mechanistic interactions in gene action
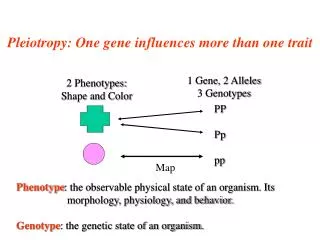
GENE INTERACTIONS • Pleiotropy • single gene affects multiple traits • Epistasis • multiple genes interact to affect a trait • multiple traits interact to produce fitness • therefore, natural selection for gene combinations
PLEIOTROPY AND EPISTASIS e.g. polymorphic Batesian mimicry (palatable mimics)
Papilio memnon, a Batesian mimic model species mimics • polymorphic Batesian mimicry • (palatable mimics): • frequency-dependent selection • for rare female-limited mimic • selection for coordinated • phenotype: gene combinations • In general, selection • is epistatic • acts on combinations of genes, • rather than single loci male is non- mimetic
How do EPISTASIS AND PLEIOTROPY affect our view of evolution? Gene interactions affect genotypic frequencies at many loci. A/a (say) controls: forewing colour pattern B/b controls: hindwing pattern AB, ab combinations favoured at expense of Ab, aB Inbreeding, selection, migration etc. cause a deviation from Hardy-Weinberg equilibrium at a single locus. Similarly, selection (also migration, drift) can cause deviation from multilocus equilibria, and lead to prevalence of particular 2-locus combinations.
HOW DO WE MEASURE DISEQUILIBRIUM? Expected gametic frequencies, if two genes are independently inherited and randomly combined, can be obtained from allelic frequencies in population: Alleles A a allele pA 1-pA freq. B pB pApB (1-pA)pB b 1-pB pA(1-pB) (1-pA)(1-pB) Sum = 1 Non-randomness of the gametic frequencies means a deviation from two locus equilibrium
D is the gametic disequilibrium coefficient, or measure of deviation from 2 locus equilibrium: Gametic random frequencies= expectation + deviation pAB= pApB+ D pAb= pA(1-pB)- D paB=(1-pA)pB - D pab=(1-pA)(1-pB) + D (Obviously, the sum pAB +pAb + paB + pab = 1 ) gametic disequilibrium,D … also known as … linkage disequilibrium. Also: D = pABpab - pAbpaB
STANDARDIZATION Frequency of gamete AB, pAB= pApB+ D D can vary between a maximum of +0.25 and a minimum of –0.25, but the range is often smaller if the frequency of alleles is not exactly 0.5. To get an idea of the fraction of maximal possible disequilibrium, D is standardized in various ways. A common way is to use the correlation coefficient: Can also useRAB2, which measures fraction of variance explained by relation between the two genes.
MORE THAN TWO LOCI Even two loci: difficult maths But real evolution: 10s or hundreds of loci affect traits. Think of three loci, A,B,C, 3 possible 2-way gametic disequilibria: DAB,DAC,DBC… and one 3-way disequilibrium, DABC (the effect of the DAB on C, of DAC on B etc.) More loci, more multi-order disequilibria! Maths complex! But, if D small, can assume few interactions between loci, and loci evolve independently; can use quantitative genetics approximations(see Kevin Fowler’s lectures next week) Rest of this lecture: pairs of loci.
FACTORS THAT CAN DECREASE D Recombination reduces disequilibrium All deviation from Hardy-Weinberg is completely lost in 1 generation of random mating Deviation from 2 locus equilibrium more persistent. Recombination at a maximum of 50% of gametes. D can therefore decline by at most 50% in each generation.
Disequilibrium actually declines by a fraction given bythe recombination rate) every generation If c= % recombination, then: Dt= Dt-1 (1 -c) after many generations (t): Dt = D0(1 -c)t
Linkage disequilibrium RAB
FACTORS THAT CAN INCREASE D A: Drift - random sampling of gametic frequencies, ~ e.g. Tightly linked markers: humans and Drosophila At tightly linked sites, loss of D slow; drift, even in very large populations can therefore have an effect. B:Selection - epistatic selection (for gene combinations) For example: mimetic butterflies, Primula flower morphs… C: Migration - mixing of populations with different frequencies
Primula veris – Cowslip – Heterostyly "Thrum" morph "Pin" morph
Primula veris – Cowslip "Thrum" morph "Pin" morph
Primula veris – Cowslip "Thrum" morph "Pin" morph
Primula veris – Cowslip "Thrum" morph "Pin" morph
"Thrum" morph "Pin" morph
Primula veris – Cowslip Supergene controls 3 major components: Dominants are all 'thrum' form. Style length: G – short style & short papillae on stigma surface Pollen size: P – 'thrum' pollen ~ 1.5x dia 'pin' pollen Anther position: A – 'thrum' anther position at mouth of corolla-tube (Matings between pin & pin and thrum & thrum are also largely incompatible). ... occasional recombinants ('homostyles') occur naturally Why are all thrum phenotypes dominant? Why does each morph only have one allele at each gene (i.e. linkage disequilibrium complete, D≈1), and not recombinant phenotypes?
Human Leucocyte Antigens (HLA): • Part of Major Histocompatibility Complex (MHC), a large complex of loci involved in the immune system. • Involved in antibody/antigen reactions, involved in recognition, presents antigen to T-cell – for lysis • Highly polymorphic, involved in immunity to disease • Probable frequency-dependent selection for rare forms • Disequilibria over 10s-100s of millions of b.p., suggesting selection for combinations of loci.
USES for gametic disequilibria 1) Studying migration or dispersal ·between populations with different gene frequencies ·between species Mixing will produce disequilibrium which will persist for some generations
2)Linkage mapping of loci when c = 0.01 or less difficult For example, genetic disease locus D and marker loci m In humans 1 million b.p. givesc≈ 1% = 1 map unit (centimorgan) Can use “association studies” or “disequilibrium mapping”, to find disease genes and markers in populations. Humans: disequilibria significant between marker loci (e.g. microsatellites, SNPs) and between markers and genetic disease loci ~ 1Mb apart, due to drift Linkage disequilibria useful for fine-scale gene mapping. D can quickly narrow search for the "candidate loci". Successfully used in recent studies. "Hapmap" project.
Linkage diseq. on human chromosome 22 Physical distance (kB)
Papilio memnon, a Batesian mimic model species mimics male is non-mimetic
3) Human mitochondrial DNA recombination? (now generally dis- believed for humans, but is probable in other species) Awadalla et al. Science 286: 2524-5
SUMMARY Gene interactions: pleiotropy, epistasis Evolution may affect frequencies of two- and more locus associations, as well as just gene frequencies. Deviation from 2-locus equilibrium is known as gametic disequilibrium or linkage disequilibrium, measured by D D is destroyed by recombination, c, so Dt= D0(1 -c)t D can increased by selection, migration, drift D is involved in maintaining 'supergenes'. D can be used in linkage mapping, studies of migration in natural populations READINGS: Futuyma: Chapter 9: 205-207, Chapter 13: 303-304 Freeman & Herron: Chapter 7