Download
1 / 20
200 likes | 424 Views
TEMA 6.- ELS SISTEMES D’ENDOMEMBRANA. En contrast amb la cèl·lula procariota, la cèl·lula eucariota conté compartiments funcionals delimitats amb membrana: els orgànuls.

E N D
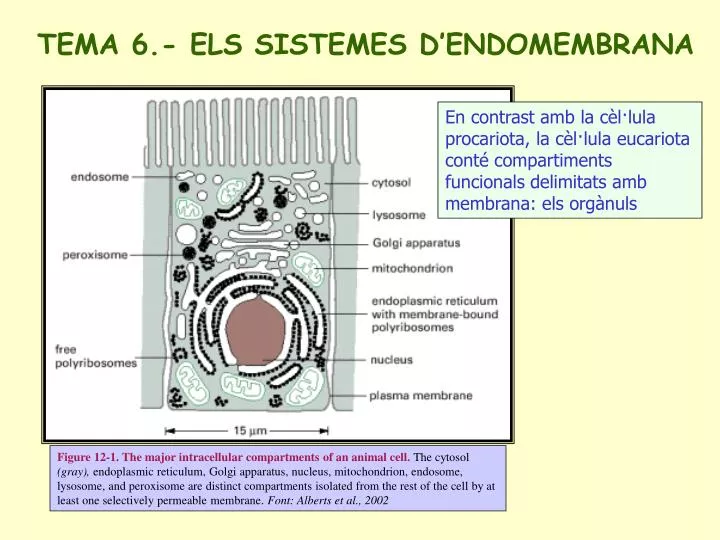
TEMA 6.- ELS SISTEMES D’ENDOMEMBRANA En contrast amb la cèl·lula procariota, la cèl·lula eucariota conté compartiments funcionals delimitats amb membrana: els orgànuls Figure 12-1. The major intracellular compartments of an animal cell. The cytosol (gray), endoplasmic reticulum, Golgi apparatus, nucleus, mitochondrion, endosome, lysosome, and peroxisome are distinct compartments isolated from the rest of the cell by at least one selectively permeable membrane. Font: Alberts et al., 2002
EL RETICLE ENDOPLÀSMIC El reticle endoplàsmic és una xarxa de túbuls i sacs que s’estén des de la membrana nuclear pel citoplasma. La membrana és contínua i suposa al voltant de la meitat de l’ àrea de membrana de la cèl·lula. És l’orgànul més gran i representa al voltant del 10% del volum de la cèlula. (A) Abundant smooth ER in a steroid-hormone-secreting cell. This electron micrograph is of a testosterone-secreting Leydig cell in the human testis. (B) A three-dimensional reconstruction of a region of smooth ER and rough ER in a liver cell. The rough ER forms oriented stacks of flattened cisternae, each having a lumenal space 20 30 nm wide. The smooth ER membrane is connected to these cisternae and forms a fine network of tubules 30 60 nm in diameter. Font: Alberts et al., 2002 Hi ha dos tipus diferents de reticle endoplasmic: el RE rugós, cobert per ribosomes, que és clau en la síntesi i el processament de proteïnes, i el RE llis, implicat en el metabolisme de lípids.
Hipòtesi sobre l’origen evolutiu del nucli, el reticle endoplàsmic i les mitocòndries The origins of mitochondria, chloroplasts, ER, and the cell nucleus can explain the topological relationships of these intra-cellular compartments in eucaryotic cells. (A) A possible pathway for the evolution of the cell nucleus and the ER. In some bacteria the single DNA molecule is attached to an invagination of the plasma membrane. Such an invagination in a very ancient procaryotic cell could have rearranged to form an envelope around the DNA, while still allowing the DNA access to the cell cytosol (as is required for DNA to direct protein synthesis). This envelope is presumed to have eventually pinched off completely from the plasma membrane, producing a nuclear compartment surrounded by a double membrane. As illustrated, the nuclear envelope is penetrated by communicating channels called nuclear pore complexes. Because it is surrounded by two membranes that are in continuity where they are penetrated by these pores, the nuclear compartment is topologically equivalent to the cytosol; in fact, during mitosis the nuclear contents mix with the cytosol. The lumen of the ER is continuous with the space between the inner and outer nuclear membranes and topologically equivalent to the extracellular space. (B) Mitochondria (and plastids) are thought to have originated when a bacterium was engulfed by a larger pre-eucaryotic cell. They retain their autonomy. This may explain why the lumens of these organelles remain isolated from the membrane traffic that interconnects the lumens of many other intracellular compartments. Font: Alberts, et al., 2002
El paper del reticle endoplàsmic rugós en la secreció de proteïnes Figure 9.2. The secretory pathway Pancreatic acinar cells, which secrete most of their newly synthesized proteins into the digestive tract, were labeled with radioactive amino acids to study the intracellular pathway taken by secreted proteins. After a short incubation with radioactive amino acids (3-minute label), autoradiography revealed that newly synthesized proteins were localized to the rough ER. Following further incubation with nonradioactive amino acids (a chase), proteins were found to move from the ER to the Golgi apparatus and then, within secretory vesicles, to the plasma membrane and cell exterior. (Font: Cooper, 2000)
Les proteïnes sintetitzades als ribosomes del REr van destinades a: • Secreció a l’espai extracel·lular • Membrana plasmàtica • Lisosomes o membrana lisosomal • Aparell de Golgi o membrana de les cisternes del Golgi • RE o membrana del RE EN CONTRAST • Les proteïnes sintetitzades als ribosomes lliures del CITOSOL van destinades a: • Quedar-se al citosol • Nucli • Mitocòndries • Peroxisomes • Cloroplasts Però, quines proteïnes són dirigides cap al reticle?
La hipòtesi de la seqüència senyal 4. Continua la traducció, i la cadena polipeptídica en creixement és translocada a través de la membrana 2. La PRS condueix el complex fins a la membrana del RE, on s’uneix al seu receptor 1. La seqüència senyal és reconeguda a l’eixida del ribosoma per la partícula de reconeixement del senyal (PRS) 3. La PRS s’allibera, el ribosoma s’uneix a un complex de translocació de membrana i la seqüència senyal s’inserta en un canal de membrana 5. La seqüència senyal és escindida per una peptidasa Cooper, 2000
La inserció en la bicapa lipídica de les proteïnes trans-membrana està dirigida pel procés de translocació al RE Figure 9.11. Insertion of a membrane protein with a cleavable signal sequence and a single stop-transfer sequence The signal sequence is cleaved as the polypeptide chain crosses the membrane, so the amino terminus of the polypeptide chain is exposed in the ER lumen. However, translocation of the polypeptide chain across the membrane is halted by a transmembrane stop-transfer sequence that closes the Sec61 translocation channel and exits the channel laterally to anchor the protein in the ER membrane. Continued translation results in a membrane-spanning protein with its carboxy terminus on the cytosolic side.Font: Cooper, 2000
Font: Cooper, 2000 Inserció en la bicapa lipídica d’una proteïna amb diversos dominis trans-membrana
El reticle endoplàsmic rugós te un paper clau en el plegament i processament de proteïnes • Escissió del pèptid senyal • Plegament – chaperones moleculars • Acoblament de proteïnes multimèriques • Formació de ponts di-sulfur (disulfur isomerasa) • Glicosilació de residus específics d’asparragina • Addició de glicolípids per ancorar-se a membrana Font: Lodish et al., 1999
La major part dels lípids de membrana es sintetitzen en el reticle endoplàsmic llis (A) Because new lipid molecules are added only to the cytosolic half of the bilayer and lipid molecules do not flip spontaneously from one monolayer to the other, a membrane-bound phospholipid translocator (called a scramblase) is required to transfer lipid molecules from the cytosolic half to the lumenal half so that the membrane grows as a bilayer. The scramblase is not specific for particular phospholipid head groups and therefore equilibrates the different phospholipids between the two monolayers. (B) Fueled by ATP hydrolysis, a head-group-specific flippase in the plasma membrane actively flips phosphatidylserine and phosphatidylethanolamine directionally from the extracellular to the cytosolic leaflet, creating the characteristically asymmetric lipid bilayer of the plasma membrane of animal cells. A scramblase is also present in plasma membranes to ensure that both monolayers remain equally populated with lipids; the continuous action of the flippase is therefore necessary to maintain the phospholipid asymmetry. Alberts et al., 2002 El paper dels translocadors de fosfolipidsen la síntesis de la bicapa lipídica ESCRAMBLASA FLIPASA
Proteïnes i lípids són exportades des del reticle endoplàsmic en vesícules de transport Vesicular transport from the ER to the Golgi Proteins and lipids are carried from the ER to the Golgi in transport vesicles that bud from the membrane of the ER and then fuse to form the vesicles and tubules of the ER-Golgi intermediate compartment (ERGIC). Lumenal ER proteins are taken up by the vesicles and released into the lumen of the Golgi. Membrane proteins maintain the same orientation in the Golgi as in the ER. Compartiment intermedi RE-Golgi (ERGIC) Proteins destined to remain in the lumen of the ER are marked by the sequence Lys-Asp-Glu-Leu (KDEL) at their carboxy terminus. These proteins are exported from the ER to the Golgi in the nonselective bulk flow of proteins through the secretory pathway, but they are recognized by a receptor in the ER-Golgi intermediate compartment (ERGIC) or the Golgi apparatus and selectively returned to the ER Font: Cooper, 2000
L’APARELL DE GOLGI Les diferents regions de l’aparell de Golgi Vesicles from the ER fuse to form the ER-Golgi intermediate compartment, and proteins from the ER are then transported to the cis Golgi network. Resident ER proteins are returned from the ER-Golgi intermediate compartment and the cis Golgi network via the recycling pathway. The medial and trans compartments of the Golgi stack correspond to the cisternae in the middle of the Golgi complex and are the sites of most protein modifications. Proteins are then carried to the trans Golgi network, where they are sorted for transport to the plasma membrane, secretion, or lysosomes. Font: Cooper, 2000.
L’aparell de Golgi al microscopi electrònic A) Three-dimensional reconstruction from electron micrographs of the Golgi apparatus in a secretory animal cell. The cis-face of the Golgi stack is that closest to the ER. Alberts et al., 2002 B) A thin-section electron micrograph emphasizing the transitional zone between the ER and the Golgi apparatus in an animal cell. Alberts et al., 2002 C) An electron micrograph of a Golgi apparatus in a plant cell (the green alga Chlamydomonas) seen in cross section. Alberts et al., 2002
La distribució de proteïnes des del trans-Golgi Proteïnes i glicolípids són transportats en vesícules des del trans Golgi a les seues destinacions finals In the absence of specific targeting signals, proteins are carried to the plasma membrane by constitutive secretion. Alternatively, proteins can be diverted from the constitutive secretion pathway and targeted to other destinations, such as lysosomes or regulated secretion from the cells.Font: Cooper, 2000
El transport de vesícules des del trans-Golgi als lisosomes Figure 13-37. The transport of newly synthesized lysosomal hydrolases to lysosomes. The precursors of lysosomal hydrolases are covalently modified by the addition of mannose 6-phosphate (M6P) groups in the cis Golgi network. They then become segregated from all other types of proteins in the trans Golgi network because adaptins in the clathrin coat bind the M6P receptors, which, in turn, bind the modified lysosomal hydrolases. The clathrin-coated vesicles produced bud off from the trans Golgi network and fuse with late endosomes. At the low pH of the late endosome, the hydrolases dissociate from the M6P receptors, and the empty receptors are recycled to the Golgi apparatus for further rounds of transport. It is not known which type of coat mediates vesicle budding in the M6P receptor recycling pathway. In the late endosomes, the phosphate is removed from the mannose sugars attached to the hydrolases, further ensuring that the hydrolases do not return to the Golgi apparatus with the receptor. Font: Alberts et al., 2002
Histochemical visualization of lysosomes.Electron micrograph of a cell stained to reveal the location of acid phosphatase, a marker enzyme for lysosomes. The precipitates are produced when tissue fixed with glutaraldehyde (to fix the enzyme in place) is incubated with a phosphatase substrate in the presence of lead ions. Two small vesicles thought to be carrying acid hydrolases from the Golgi apparatus are indicated by red arrows in the top panel. (Courtesy of Daniel S. Friend.) Font: Alberts et al., 2002 LISOSOMES Three pathways to degradation in lysosomes. (A) Each pathway leads to the intracellular digestion of materials derived from a different source. (B) An electron micrograph of an autophagosome containing a mitochondrion and a peroxisome. (B, courtesy of Daniel S. Friend, from D.W. Fawcett, A Textbook of Histology, 12th edn. New York: Chapman and Hall, 1994.) Font: Alberts et al., 2002
LES PROTEÏNES DIRECTORES DE LA FORMACIÓ DE VESÍCULES Utilization of different coats in vesicular traffic. Different coat proteins select different cargo and shape the transport vesicles that mediate the various steps in the biosynthetic- secretory and endocytic pathways. When the same coats function in different places in the cell, they can incorporate different coat protein subunits that modify their properties (not shown). Many differentiated cells have additional pathways beside those shown in this figure, including a sorting pathway from the trans Golgi network to the apical surface in polarized cells and a specialized recycling pathway for proteins of synaptic vesicles in the synapses of neurons. Font: Alberts et al., 2002
Les vesícules revestides de clatrina Incorporation of lysosomal proteins into clathrin-coated vesicles Proteins targeted for lysosomes are marked by mannose-6-phosphates, which bind to mannose-6-phosphate receptors in the trans Golgi network. The mannose-6-phosphate receptors serve as binding sites for cytosolic adaptor proteins, which in turn bind clathrin. Clathrins consist of three protein chains that associate with each other to form a basketlike lattice that distorts the membrane and drives vesicle budding. Font: Cooper, 2000
Les vesícules revestides de COPs Role of ARF in the formation of COP-coated vesicles ARF alternates between GTP-bound and GDP-bound states. When bound to GDP, ARF associates with the membrane of the trans Golgi network, where guanine nucleotide exchange factors (GEF) promote the exchange of the bound GDP for GTP. In its GTP-bound state, ARF promotes the binding of COPI coat protein (coatomer), leading to vesicle budding. Hydrolysis of the bound GTP then converts ARF to the GDP-bound state, leading to disassembly of the vesicle coat prior to fusion with the target membrane. Font: Cooper, 2000
La fusió de vesícules Vesicle fusion Vesicle fusion is mediated by binding between specific pairs of v-SNAREs and t-SNAREs on the vesicle and target membranes, respectively. Rab GTP-binding proteins are required to facilitate formation of v-SNARE/t-SNARE complexes. Following membrane fusion, the NSF/SNAP proteins disassemble the SNARE complex. Font: Cooper, 2000
