Download
1 / 28
280 likes | 562 Views
Potencjał czynnościowy - propagacja.

E N D
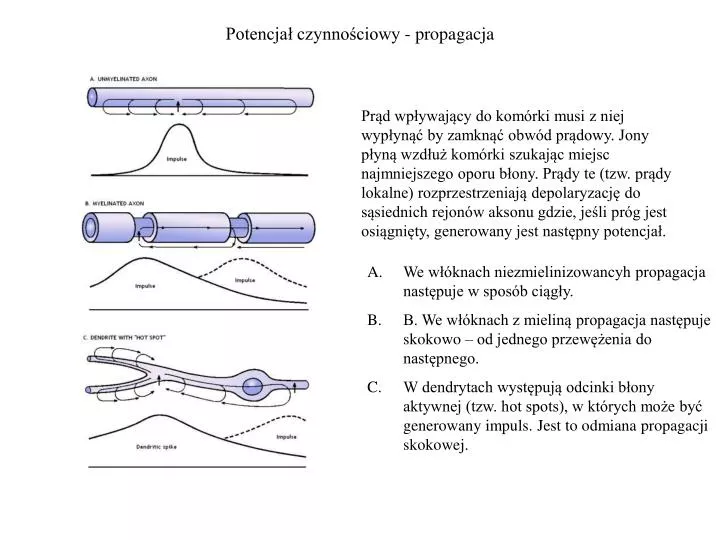
Potencjał czynnościowy - propagacja Prąd wpływający do komórki musi z niej wypłynąć by zamknąć obwód prądowy. Jony płyną wzdłuż komórki szukając miejsc najmniejszego oporu błony. Prądy te (tzw. prądy lokalne) rozprzestrzeniają depolaryzację do sąsiednich rejonów aksonu gdzie, jeśli próg jest osiągnięty, generowany jest następny potencjał. • We włóknach niezmielinizowancyh propagacja następuje w sposób ciągły. • B. We włóknach z mieliną propagacja następuje skokowo – od jednego przewężenia do następnego. • W dendrytach występują odcinki błony aktywnej (tzw. hot spots), w których może być generowany impuls. Jest to odmiana propagacji skokowej.
Skokowa propagacja potencjału czynnościowego W obszarach zmielinizowanych błona ma duży opór oraz małą pojemność, co powoduje, że prąd płynie pomiędzy przewężeniami Ranviera. W przewężeniach znajduje się duża liczba kanałów Na+. Kanały K+ znajdują się pod mieliną, pomiędzy przewężeniami.
Teoria kablowa (cable theory) – propagacja potencjału czynnościowego Aby opisać propagację potencjału czynnościowego należy rozważyć teorię kablową opisującą rozchodzenie się jonów wzdłuż aksonu. W teorii kablowej aksony i dendryty są traktowane jako cylindry złożone z pojemności cm i oporu rm połączonych równolegle. Wzdłuż włókna występuje opór ri połączony szeregowo. ri rm, cm własności membrany parametry kabla - wewnątrzkomórkowy opór właściwy ( W*cm) - oporność osiowa (W /cm ) - opór właściwy błony (W *cm2) - oporność błony (W * cm ) - pojemność właściwa błony (F/cm2 ) - pojemność błony (F/cm ) Parametry membrany o długości l, przekroju A = pa2 i powierzchni S = 2pal : Ri, total=Ri*l/A Rm, total=Rm/S Cm, total=Cm*S
Teoria kablowa (cable theory) - cd Prąd przez błonę: Prawo Ohma: Zanik ii wraz z odległością (wyciek przez rm i cm): Potencjał czynnościowy propaguje się ze stałą prędkością. Stosujemy więc równanie falowe: Dostajemy gdzie, - prędkość propagacji
Teoria kablowa (cable theory) - cd Po rozwiązaniu równania dostajemy: dla K = 10.47 m/s – wyznaczona doświadczalnie q= 18.8 m/s qexp= 21.2 m/s
Dwa mity Neuron Impulsy elektryczne Impulsy elektryczne Komunikacja komórka grzyba komórka jajowa strunowca komórka skóry żaby pień dyni komórka przysadki mógowej szczura Komórki Rosiczki generują impulsy elektryczne podczas zamykania listków komórka pierwotniaka komórka trzustki szczura
Synapsa Sir Charles Sherrington, 1897, Podręcznik fizjologii <gr. sýnapsis połączenie>
Synapsy chemiczne, elektryczne i nie tylko... Metody komunikacji w układzie nerwowym: Przekaźnictwo objętościowe (volume transmission) Zbliżenia membrannowe (membrane juxtapositions) Synapsy elektryczne Synapsy chemiczne Zbliżenie membrannowe we włóknach nizemielinizowanych, umożliwiające komunikacje poprzez jony K+ lub przepływ prądu (--). Rodzaje połączeń membrannowych
Synapsy chemiczne, elektryczne Distinguishing Properties of Electrical and Chemical Synapses Cytoplasmic continuity between pre- and postsynaptic cells Ultrastructural components Synaptic delay Type of synapse Distance between pre- and postsynaptic cell membranes Agent of transmission Direction of transmission Yes Gap-junction channels Virtually absent Electrical 3.5 nm Ion current Usually bidirectional No Presynaptic vesicles and active zones; postsynaptic receptors Significant: at least 0.3 ms, usually 1-5 ms or longer Chemical 20-40 nm Chemical transmitter Unidirectional
Synapsy elektryczne A. W synapsie elektrycznej dwie komórki są połączone kanałami szczelinowymi (gap-junction channels). Kanały te umożliwiają bezpośredni przepływ jonów pomiędzy dwoma komórkami. Dodatkową ułatwieniem komunikacji jest zawężenie przestrzeni zewnątrzkomórkowej z 20 nm do 3.5 nm w złączu szczelinowym (gap junction). Mikrografia elektronowa połączenia szczelinowego. Macierz kanałów wyizolowana z błony wątroby szczura. Każdy kanał ma strukturę hexagonalną. Powiększenie: X 307 800 B. Każdy półkanał (connexon) składa się z sześciu identycznych podzespołów (connexin). C. Podzespoły są ułożone tak, że tworzą por pośrodku kanału. Por jest otwarty gdy podzespoły są skręcone względem podstawy. Na otwarcie lub zamknięcie poru może wpływać poziom pH i stężenie Ca2+ w komórce. Synapsy elektryczne mogą mieć również napięciowozależne bramki oraz reagować na różne neuroprzekaźniki. • Główne cechy przekaźnictwa elektrycznego: • duża prędkość • wierność przekazu (bez zniekształcenia) • działanie dwukierunkowe • Zastosowanie: • szybkie działanie (np. odruch ucieczki) • synchroniczne działanie dużych grup neuronów • komunikacja w komórkach glejowych
Synapsy elektryczne u Aplysii E. Kandel z Aplysią Stymulacja elektryczna ogona powoduje synchroniczne wyładowanie w trzech motoneuronach. 1. Gdy motoneurony znajdują się w spoczynku stymulacja wywołuje identyczną salwę potencjałów czynnościowych w trzech komórkach czego wynikiem jest wypuszczenie atramentu. 2. W stanie hiperpolaryzacji, odpowiedź obronna jest zablokowana.
Synapsa chemiczna • Depolaryzacja błony presnaptycznej. • Aktywacja napięciowozależnych kanałów Ca2+ • Fuzja błony pęcherzyka synaptycznego z błoną presynaptyczną • Uwalnianie neurotransmitera (egzocytoza) • Dyfuzja neurotransmitera do szczeliny synaptycznej • Przyłączenie neurotrasmitera do receptora i bramkowanie kanałów jonowych • Odnowa pęcherzyka synaptycznego –endocytoza • Inaktywacja neurotransmitera
Różnorodność połączeń synaptycznych Znakowane fluorescencyjnie synapsy (pomarańczowe punkty) w komórce hipokamapa. Mikrografia elektronowa szczeliny synaptycznej. Duże owalne struktury – mitochondria, małe okrągłe struktury - pęcherzyki synaptyczne. Rozmyte zgrubienia (strzałki) – strefy aktywne – obszary zawierające dużą gęstość pęcherzyków synaptycznych i receptorów.
Złącze nerwowo – mięśniowe (neuromuscular junction) Znakowane włókna mięśniowe i rozgałęzienia aksonu motoneuronu dochodzące do płytek końcowych. Mięsień jest unerwiany przez motoneuron. Rozgałęzienia motoneuronu tworzą połączenia synaptyczne z pojedynczymi włóknami mięśniowymi. Każde złącze, tzw. płytka końcowa, składa się z: zakończenia włókna nerwowego skąd uwalniana jest acetylocholina (ACh), szczeliny synaptycznej, obszaru postsynaptycznego zawierającego receptory acetylocholiny, otoczki glejowej. Zdjęcie z mikroskopu elektronowego ukazujące przekrój złącza nerwowo mięśniowego. T – zakończenie aksonu, M – włókno mięśniowe. Skala 0.3 mm.
Złącze nerwowo – mięśniowe – potencjały w płytce końcowej Własności synapsy na przykładzie złącza nerwowo – mięśniowego. A. Zapisy wewnątrzkomórkowe potencjałów w płytce końcowej prowadzące do potencjału czynnościowego w komórce mięśnia; z lewej - układ eksperymentalny. B. Sumowanie miniaturowych potencjałów w płytce końcowej (MEPP). Pojedynczy MEPP wywołane jest uwolnieniem porcji jednostkowej Ach (~10000 cząsteczek). Uwolnienie wielu porcji jednostkowych w tym samym czasie powoduje sumowanie się potencjałów MEPP. C. Szum wywołany jonoforezą (wprowadzaniem pod wpływem pola elektrycznego) pojedynczych cząsteczek ACh. D. Zapis patch – clamp pokazujący prądy płynące przez pojedynczy kanał AChR.
Hipoteza kwantowa (del Castillo, Katz, Martin) Uwalnianie neurotransmitera ma naturę probabilistyczną, a nie deterministyczną m = np m – zawartość kwantowa tj. ilość uwolnionych kwantów (porcji jednostkowych) n – ilość dostępnych kwantów p – prawdopodobieństwo uwolnienia n ~ 1000 (ilość stref aktywnych) m ~ 100-200 p ~ 0.1 – 0.2 Kwantowe uwalnianie neurotransmitera stosuje się do większości synaps chemicznych. Proces (prawdopodobieństwo) uwalniania jest kontrolowany depolaryzacją błony zakończenia synaptycznego.
Kanał jonowy w płytce końcowej jest przepuszczalny dla Na+ i K+ Prąd synaptyczny płytki końcowej: IEPSP =gEPSP(V – VEPSP) gEPSP – przewodnictwo kanału ACh, V – potencjał błonowy, VEPSP – potencjał równowagowy dla jonów płynących przez kanał ACh. Prądy jonowe tworzące potencjał płytki końcowej można badać mierząc potencjał równowagowy prądu płytki końcowej. Napięcie na błonie komórki mięśnia jest ustalane na różnych wartościach i mierzony jest prąd synaptyczny podczas stymulacji mięśnia. A.Gdyby jedynie wpływ Na+ powodował IEPSP, potencjał odwrócenia wystąpiłby przy potencjale równowagowym dla sodu +55mV. B. Prąd płytki końcowej odwraca się przy 0 mV gdyż kanał przewodzi jony Na+ i K+. Prąd wypadkowy jest sumą wpływów i wypływów Na+ i K+. Przy potencjale odwrócenia wypływ Na+ jest równoważony wypływem K+.
Badanie synaps plaster hipokampa komora in vitro hodowane kultury bakterii mózg in toto
Hipokamp Ewolucja hipokampa na przykładzie niższego kręgowca (rekin), ssaka (szczur) i człowieka. Konik morski (Hippocampus) Organizacja hipokampa. A Kora śródwęchowa (entorhinal cortex), B. podkładka (subiculum), C. Hipokamp, D. Zakręt zębaty (dentate gyrus), E. Sklepienie (fornix), F. Włókna do kory śródwęchowej, G. Koryto (alveus), H. Kora okołokomorowa (periventricular grey matter).
Synapsy pobudzające Dwa rodzaje receptorów glutaminianowych: AMPA: α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid NMDA: N-methyl-D-aspartate Odpowiedzi receptora AMPA i NMDA przy różnych wartościach potencjału. Separacja aktywności dwóch rodzajów receptora jest uzyskana selektywną blokadą receptora NMDA przez APV.
Własności receptora NMDA bez Mg2+ z Mg2+ - Kanał NMDA jest napięciowozależny ze względu na napięciowozależną blokadę Mg+
Salwy epileptyczne in vitro RD Traub, JG Jefferys and MA Whittington. Enhanced NMDA conductance can account for epileptiform activity induced by low Mg2+ in the rat hippocampal slice, The Journal of Physiology, 1994, 478 (3) 379-393.
Synapsy hamujące Dwa rodzaje receptorów GABA (γ-aminobutyric acid) GABAA GABAB Odpowiedzi receptora AMPA, GABAA i GABAB. Potencjały odwrócenia –90 mV (GABAB) i – 70 mv (GABAA) wskazują udział jonów K+ i Cl-.
Receptory jonotropowe i metabotropowe Neurotrasmitery mogą działać bezpośrednio i pośrednio na kanały jonowe regulujące przepływ jonów. Bezpośrednie bramkowanie odbywa się w receptorach jonotropowych. Bramkowanie pośrednie odbywa się poprzez aktywację receptorów metabotropowych. Receptory metabotropowe aktywują białko G, które aktywuje kaskadę procesów modulujących aktywność kanału jonowego. GABAA: neuroprzekaźnik -> otwarcie kanału Cl- GABAB : neuroprzekaźnik -> aktywacja białka G -> podwyższenie poziomu cAMP (cykliczny adenozynomonofosforan) ->otwarcie kanału K+
Receptory jonotropowe i metabotropowe Receptory jonotropowe powodują szybką i krótkotrwałą odpowiedź synaptyczną. Występują w obwodach kontrolujących szybkie zachowania. Receptory metabotropowe dają odpowiedź wolniejszą i dłuższą. Modulują zachowanie zmieniając pobudliwość neuronów i siłę połączenia synaptycznego.
Hamowanie bocznikujące (shunting inhibition) Efekt bocznikowania.Gdy komórka dostaje wejście pobudzające i hamujące, kanały otwarte przez hamowanie bocznikują (wypuszczają) prąd pobudzający zmniejszając pobudzający potencjał synaptyczny. Bocznice w Holandii
Narkotyki a neuroprzekaźnictwo Działanie różnych leków (nasenne, antydepresyjne) oraz narkotyków polega na ingerencji w neuroprzekaźnictwo. Np. marijuana i heroina aktywuje neurony ponieważ ich chemiczna struktura jest podobna do naturalnych neuroprzekaźników. Pomimo ze skutecznie ‘podszywają’ się one pod neuroprzekaźniki, aktywacja neuronow nie jest taka sama i prowadzi do zmienionej aktywnosci sieci. amfetamina, kokaina powoduje zwieksza stezenie neuroprzekaźników poprzez ich wydzielanie w wiekszych niż normalnie ilosciach lub poprzez blokade ich wychwytu zwrotnego. Wiekszosc narkotykow np. kokaina, amfetamina wplywa na uklad nagrody zwiazany z dopamina (DA). DA jest obecna w rejonach mozgu odpowiedzialnych m.in. za motywacje, uczucia i odczuwanie przyjemnosci. Leki psychiatryczne wyrównują niedobór lub nadmiar naturalnych neuroprzekaźników. Odbywa się to np. poprzez blokowanie wychwytu zwrotnego (np. serotoniny 5-HT) przez leki andytdepresyjne (Prozac, Paxil, Zoloft). Inne leki ‘podszywają’ się lub blokuja enzymy rozkładajace neuroprzekaźniki. Leki nasenne i uspokajające zwiększają aktywność receptorów hamujących GABA, hamując poziom aktywności człowieka.



