Download
1 / 27
300 likes | 681 Views
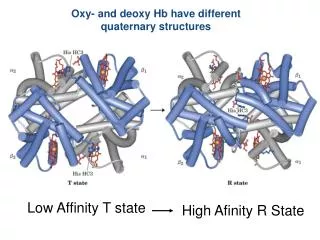
Oxy- and deoxy Hb have different quaternary structures. Low Affinity T state. High Afinity R State. Hb is regulated by H + and CO 2. lungs. More O 2 released as the pH is lowered. tissue. The Bohr effect : the effect of pH and CO 2 on the binding and release of oxygen to Hb.

E N D
Oxy- and deoxy Hb have different quaternary structures Low Affinity T state High Afinity R State
Hb is regulated by H+ and CO2 lungs More O2 released as the pH is lowered tissue The Bohr effect: the effect of pH and CO2 on the binding and release of oxygen to Hb. Lower pH (higher H+) stabilizes the T-state
Hb binds oxygen cooperatively Hb sigmoidal curve is a composite of hyperbolic T and R state curves Deoxy Hb is in T state and has reduced O2 affinity. Hb stays in T state until 1 or 2 dioxygens are bound Then the quaternary structures snaps to R, changing the conformations of all 4 subunits, whether or not they are bound with dioxygen R State T state
Ion pairs that stabilize the T state of deoxyhemoglobin must break to form R state
Protonation of His HC3 in T state is a major contributor to Bohr effect
CO2 (high in tissues) is also carried by hemoglobin This reaction produces protons and additional salt bridges stabilize the T state.
BPG binding stabilizes the T-state of deoxyHb BPG binds in central cavity • BPG preferentially binds to deoxy state (T state) • BPG shifts the T R equilibrium towards the T state • hence lower affinity for dioxygen
Free energy change is dependent on concentration DG Actual free energy change under specified conditions, including concentration of reactants and products DG0 Standard Free energy change, all reactants and products in their standard states, i.e. 1 M concentration. Unfortunately, if [H+] = 1 M, pH = 0, which is not consistent with biochemical processes. DG0' Standard Free Energy change for the biochemical standard state, all reactants and products at 1 M except [H+] = [OH-] = 10-7 M, which allows pH = 7.
Consider a reaction: A + B C Go = free energy change of the reaction under standard conditions • G = Go + RT ln Keq • G = Go + RT ln([C]/[A][B]) At equilibrium,G = 0 Therefore, Go = - RT ln Keq = - RT ln ([C]/[A][B]) Thus the equilibrium constant can be calculated from standard free energy data and vice versa.
Standard state conditions Reactions 1-10 are glucose to pyruvate and reaction 11 is pyruvate to lactate. At concentrations in RBC From Lehninger Principles of Biochemistry
Under physiological conditions except for 1,3 & 4 all reaction have DG values closer to zero Standard free energy changes for TCA cycle DG' = -32.2 kJ/mol DG' = 29.7 kJ/mol DG' = 13.3 kJ/mol DG' = -3.8 kJ/mol DG' = -20.9 kJ/mol DG' = 0 kJ/mol DG' = -33.5 kJ/mol DG' = -2.9 kJ/mol From Lehninger Principles of Biochemistry
An unfavorable reaction can proceed spontaneously if it is coupled to an energetically favorable reaction A B + X DG = + 50 kJ/mol X Y + Z DG = -100 kJ/mol Overall reaction: A B + Y + Z DG = - 50 kJ/mol
Values from human erythrocytes PEP + H2O Pyruvate + PiDG = -78 kJ/mol ADP + Pi ATP + H2O DG = 55 kJ/mol PEP + ADP Pyruvate + ATP Total DG = -23 kJ/mol
Classes of Inhibition Two real, one hypothetical • Competitive inhibition - inhibitor (I) binds only to E, not to ES • Uncompetitive inhibition - inhibitor (I) binds only to ES, not to E. This is a hypothetical case that has never been documented for a real enzyme, but which makes a useful contrast to competitive inhibition • Noncompetitive (mixed) inhibition - inhibitor (I) binds to E and to ES
Inhibitor (I) binds only to E, not to ES Inhibitor (I) binds only to ES, not to E. This is a hypothetical case that has never been documented for a real enzyme, but which makes a useful contrast to competitive inhibition Enzyme Inhibition Inhibitor (I) binds to E and to ES. From Lehninger Principles of Biochemistry
CompetitiveUncompetitiveNoncompetitive InhibitionInhibition(Mixed) Inhibition Kmchanges while Vmax does not Km and Vmax both change Km and Vmax both change From Lehninger Principles of Biochemistry
Regulation of enzymatic activity • Two ways that this may occur: • Control of enzyme availability • Depends on rate of enzyme synthesis & degradation • Control of enzyme activity • Enzyme-substrate binding affinity may vary with binding of small molecules called allosteric effectors (ex: BPG for Hb) • Allosteric mechanisms can cause large changes in enzymatic activity
1. Allosteric enzymes 2. Regulation by covalent modification Regulatory Enzymes important in controlling flux through metabolic pathways From Lehninger Principles of Biochemistry
Regulation by Feedback Inhibition Conversion of L-threonine to L-isoleucine catalyzed by a sequence five enzymes, E1-E5 L-isoleucine is an inhibitory allosteric modulator of E1 From Lehninger Principles of Biochemistry
Example for Control of enzyme activity Allosteric enzymes AMP F-2,6-bis P Binds ATP only in T state Regulatory site ATP PFK subunit Citrate Active site Can bind ATP in T or R state ATP Substrates F-6-P Preferentially binds to the R state inhibition activation • ATP inhibits • AMP reverses inhibition • Citrate is also an allosteric inhibitor • Fructose-2,6-bisphosphate is allosteric activator
From Lehninger Principles of Biochemistry
Model of the E. coli F1F0–ATPase Page 832
Energy-dependent binding change mechanism for ATP synthesis by proton-translocating ATP synthase