Download

1 / 19
200 likes | 408 Views
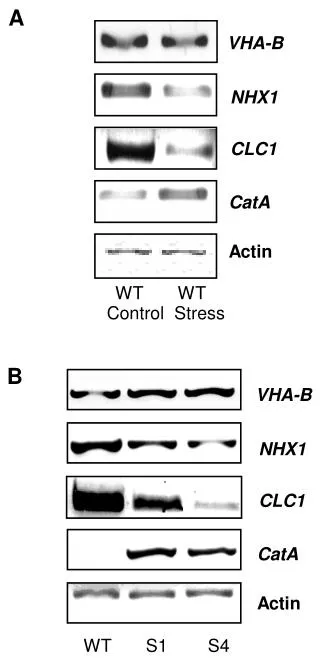
APBs involved in regulating actin dynamics (Cont.). Lodish 5th Ed. Chapter 19, p786-791. 2. How high rates of actin polymerization are maintained at the protruding edge? Thymosin ï¢-4 Profilin Arp2/3 Capping Severing . How is Arp2/3 activated. N-terminus. C-terminus.

E N D
APBs involved in regulating actin dynamics(Cont.) Lodish 5th Ed. Chapter 19, p786-791 • 2. How high rates of actin polymerization are maintained at the protruding edge? • Thymosin -4 • Profilin • Arp2/3 • Capping • Severing
How is Arp2/3 activated N-terminus C-terminus • Wiskott-Aldrich Syndrome protein – implicated in immune dysfunction • Scar/WAVE = WASp-related proteins • Purified ARP2/3 complex has low nucleating acitivity • Enhanced when WASp added • Enhanced further when pre-incubated w. F-actin • VCA (verprolin cofilin acidic) • Is proline rich, VCA domain essential for nucleation • Verprolin binds an actin monomer, A binds ARP2/3 • F-actin enhances its binding to ARP2/3
Activation of WASp • Is auto inhibited until activation • WASp is activated when the proline rich and GBD domain binds PIP2 and active Cdc42 (small GTPase) • Further activation = ATP hydrolysis within ARP1 or ARP2 • New filaments grow from the side of pre-existent filaments • Growth stops when + end is capped
Treadmilling in vitro is much slower than in vivo • Treadmilling rate in vitro is limited by thedepolym.rate (-end) = 0.2 monomers/sec • Resulting in a max growth rate at +end of ~ 0.05 um/min. • BUT some cells can move = >10 um/min i.e. x200 greater • To achieve rates of protrusion observed in vivo - need to supply + end with 20-200 subunits/second ! • Capping proteins • 1. Generate more + ends • 2. Increase size of G-actin pool • 3. Restrict polymerization to where it’s needed Severing proteins • Increase numbers of + and – ends • Increase G-actin pool
Table 19-2 Actin binding proteins that cap and sever F-actin • Capping proteins inhibit addition or loss of G-actin from the end (+ or -) of the actin filament • Actin filaments in muscle are very stable - capped by capZ at + end and tropomodulin at - end
The dynamics of actin filaments is greatly affected by ABPs that bind to filament ends • Filaments that are capped at the + end are less dynamic and more stable than uncapped filaments • Low concentrations of these ABPs (relative to actin monomers) are sufficient • In non muscle cells capping is regulated by PIP2 • Decrease in PIP2 (when cleaved) increases capping • An increase in PIP2 (shortly after initial signal) leads to uncapping
Severing proteins regulate the length and dynamics of actin filaments • Increase actin filament dynamics by generating a large number of shorter filaments • Gelsolin can: • inc. number of growing +ends • Inc. depolymerization from the -end • Low stoichiometry • e.g. 1 gelsolin molecule for every 400 actin monomers (in a filament) is sufficient!
Mechanism of severing • Severing proteins, e.g. gelsolin are activated in by an increase in intracellular calcium following a signaling event. • After Ca2+ inc. and severing F-actin, gelsolin caps the + end • Shortly after this, PIP2 increases and gelsolin dissociates from the + end allowing polymerization
Cytoplasmic streaming Amoeba proteus Gelsolin regulates the biomechanical properties of actin gels • Gelsolin solates actin gels allowing cytoplasmic streaming • Cytoplasm “gelates” or solidifies at the tip • Filamin and -actinin are important for gelation • Abundant =1% tot. protein in cell • Very sensitive to increases in [Ca2+]i (10-7M) during signaling • Cytoplasmic extracts in amoeba, leukocytes, fibroblasts can still stream
The stability of actin filaments is regulated by ABPs that bind to their sides • Need to be present in high concentractions e.g. • Tropomyosin binds to the sides of 7 adjacent actin monomers • Tropomyosin stabilizes the actin filaments and excludes other ABP’s from binding, therefore its activity must be regulated • In muscle, the position of tropomyosin is altered to expose myosin II binding sites, only when contraction is triggered
Cofilin de-stabilizes actin filaments • It causes the actin filaments to twist more tightly, and so weakens interactions between neighboring actin subunits • Cofilin binds preferentially to ADP actin, so its activity is restricted to “older” regions of F-actin (closer to -end). • Cofilin increases the rate of depolymerization from the - end of F-actin by 25 times
Cofilin Actin Merge Cofilin is located ~1-2um behind the leading edge Distribution of cofilin in a fish keratocyte Svitkina and Borisy, 1999
Capping and severing work synergistically to increase the rate of treadmilling The funneled treadmilling process(Carlier and Pantaloni, 1997) sustains an efficient, rapid, rate of actin polymerization Capping ADF/cofilins increase disassembly (x25) from - ends Regulated capping of +ends increases the pool of ATP-G actin Nucleotide exchange Un-capping The rate of polymerization is increased ADF/cofilins increase the ATP G-actin pool Fast + end assembly of a few uncapped filaments
The combined effect of capping, severing and depolymerizing F-actin is important for sustaining rapid protrusion From: “Control of actin dynamics” Carlier 1998
The combined effect of capping, severing and depolymerizing F-actin is important for sustaining rapid protrusion (For notes) From: “Control of actin dynamics” Carlier 1998
Treadmilling occurs within the whole population of F-actin • FACT: In a steadily migrating cell lamellipodium width is ~ constant, - due to balance between assembly at the front and disassembly further back
-actinin Arp2/3 ABP-120 Location of actin-binding proteins reflects their function At front edge ~4um behind front edge