Download
1 / 58
580 likes | 914 Views
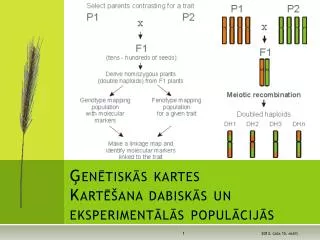
Ģenētiskās kartes Kartēšana dabiskās un eksperimentālās populācijās . Mācību plāns un lekciju saraksts . Kas notiek bivalentu veidošanās laikā? . Sinaptonemālais komplekss . Nehomologu (atšķirīgu) hromosomu bivalenti.

E N D
Ģenētiskās kartesKartēšana dabiskās un eksperimentālās populācijās
Nehomologu (atšķirīgu) hromosomu bivalenti • Piemēram, X un Y hromosomu pāris vīrišķajā mejozē, vai ģenētiski atšķirīgas homologās hromosomas • Nehomologie rajoni neveido sinaptonemālo kompleksu un krustmija nenotiek
Rekombinācijas kartes un fiziskās kartes attiecība • Zinot genoma fiziskos izmērus nukleotīdos un ģenētisko karšu izmērus centimorganīdās ir iespējams izrēķināt vidējo fiziskās un ģenētiskās kartes izmēru attiecību • Piemēram, miežu ģenētiskās kartes ir caurmērā 1250 cM lielas, bet genoma izmēri 5 x 109bp • Vidējā fiziskā un ģenētiskā attāluma attiecība ir 4 x 106bpcM-1
Rekombinācijas karstie punkti • Rekombinācija genomā nenotiek vienmērīgi – pastāv karstie un aukstie rekombinācijas punkti • Fuetal. (2002) Recombination rates between adjacent genic and retrotransposon regions in maize vary by 2 orders of magnitude. PNAS, 99: 1082 • Gēnus saturošie genoma rajoni (gēnu salas) bieži ir rekombinācijas karstie punkti
Kvalitatīvās un kvantitatīvās pazīmes • Kvalitatīvās pazīmes – diskrētas pazīmes, kuru iedzimtība atbilst Mendeļa likumam par viena gēna alēļu skaldīšanos • Kvantitatīvās pazīmes – izmērāmas pazīmes • Izšķir vairākus kvantitatīvo pazīmju veidus: • - nepārtrauktās (continuous) pazīmes (auga augstums); • - integrālās (meristic) pazīmes (vārpu skaits); • Kvantitatīvas pazīmes var noteikt viens vai vairāki gēni, tām raksturīga atkarība no apkārtējās vides ietekmes
Iedomāta miežu DH kartēšanas populācija. Fenotips – augu augstums (nepārtraukta pazīme) Nosaka fenotipu. Nosaka molekulāro marķieru genotipus Kartē QTL noteiktā molekulāro marķieru intervālā izmantojot daudzfaktoru regresijas analīzi
Iedomāta miežu DH kartēšanas populācija. Fenotips – ražība izteikta centneros no hektāra (nepārtraukta pazīme). Parādīta transgresīvā skaldīšanās
Rekombinantāsintrogresijas līnijas • Sakrusto divus dažādus vecākaugus (P1 x P2) • Veic secīgu pēcnācēju atkrustošanu ar vienu no vecākaugiem, piemēram, P2 • Iegūstrekombinantāsintrogresijas līnijas, kurās P1 genoma fragmenti atrodas P2 vecākaugagenomā • Genotipēintrogresijas līnijas ar molekulārajiem marķieriem un vienlaicīgi nosaka introgresijas līniju fenotipu • Identificē P1 genoma fragmentus, kas satur noteiktas pazīmes nosakošus QTL
Kartēšana dabiskajās populācijās • Kartēšana eksperimentālajās populācijās pamatojas uz rekombināciju un gēnu saistību hromosomā • Kartēšana dabiskajās populācijas pamatojas uz nelīdzsvaroto saistību starp dažādu gēnu alēlēm populācijā
Nelīdzsvarotā saistība • LD – allēļu asociācija starp dažādiem lokusiem. Situācija, kad divas vai vairākas allēles ir saistītas biežāk nekā to paredz populāciju ģenētikas likumi • Korelācija starp dažādiem polimorfismiem (piemēram, SNP) ir atkarīga no to izcelšanās (mutācijas) vēstures un rekombinācijas, kas ir notikusi starp dažādajiem polimorfismiem • Lielā populācijā, kurā visiem indivīdiem ir vienādas iespējas pāroties savā starpā, kurā visi lokusi skaldās neatkarīgi, kurā nenotiek izlase, mutācija un migrācija visi lokusi būs līdzsvarotā saistībā • Saistība, izlase un populāciju sajaukšanās (admixture) palielinās nelīdzsvaroto saistību
Atšķirība starp nelīdzsvaroto saistību un saistību • Saistība attiecas uz noteiku alēļu korelāciju gamētās, ko izraisa to fiziska saistība vienā hromosomā • Nelīdzsvarotā saistība attiecas uz alēļu korelāciju populācijās • Taču jāņem vērā, ka starp fiziski cieši saistītiem polimorfismiem pastāvēs arī augsts nelīdzsvarotās saistības līmenis, jo rekombinācija starp tuvu novietotiem lokusiem būs reta. Tuvu novietotus polimorfismus izlase un ģenētiskais dreifs ietekmēs līdzīgi
Izmanto esošās šķirnes, kurās potenciāli būs daudz lielāka fenotipiskā daudzveidība Izmanto šķirņu veidošanas gaitā notikušo rekombināciju Izmanto esošos fenotipiskos datus no šķirņu veidošanas laikā veiktajām pārbaudēm Problēmas var radīt populāciju struktūra Pazīmju kartēšana – asociāciju ģenētika
Kartēšanas populāciju grafisko genotipu salīdzinājums • Piemēram, DH populācijā analizēta tiek tikai viena mejoze, kuras laikā notikušas nedaudzas (1 – 3) krustmijas) • Dabiskās populācijās redzamais marķieru genotipu sadalījums ir daudzu mejožu un krustmiju rezultāts, kuru laikā rekombinācijas ceļā ir radušies radušies unikāli haplotipi
Asociāciju ģenētika testē vai pastāv korelācija starp allēļu frekvencēm marķieru lokusos un fenotipa (pazīmes, slimības, u.tmldz.) frekvencēm. Pozitīvas asociācijas iespējamas, ja a) marķieris nosaka konkrēto pazīmi; vai arī b) marķieris atrodas nelīdzsvarotā saistībā ar gēnu, kas kodē pazīmi Visa genoma asociācijas pētījumu iespēja ir atkarīga no marķieru daudzuma, kas jāpārbauda, lai identificētu asociācijas Marķieru daudzums ir atkarīgs no LD bloku izmēriem genomā. Jo lielāki LD bloki, jo mazāks skaits marķieru nepieciešams, bet vienlaicīgi samazinās kartēšanas izšķirtspēja Lander and Schork (1994) Science 265:2037 Asociāciju ģenētika
Nelīdzsvarotās saistības rašanās Ardlie et al. (2002) Nat Rev Genet, 3:299
Nelīdzsvarotā saistība Ardlie et al. (2002) Nat Rev Genet, 3:299
LD aprēķini – r2 • Divi lokusi, katrā pa divām alēlēm, Aa un B b • Alēles frekvences populācijā izsaka kā pA, pa, pB, pb • Haplotipu frekvences izsaka kā pAB, paB, paB, pab • Atšķirība starp novērotajām un sagaidāmajām haplotipu frekvencēm ir Dab= (pAB – pApB) • Tad: • r2 var uzskatīt par korelācijas koeficientu
LD aprēķini – r2 • Flint – Garciaetal. (2003) Structureoflinkagedisequilibriuminplants. Annu RevPlantBiol, 54: 357 • TASSEL (http://www.maizegenetics.net/) – Trait analysis by association, evolution and linkage • Bradburyetal. (2007) TASSELL Softwareforassociationmappingofcomplextraitsindiversesamples. Bioinformatics ,23: 2633 • r2 var uzskatīt par korelācijas koeficientu, taču r2 = 1 tikai tad, ja alēļu frekvences abos lokusos ir identiskas • r2 atkarīgs gan no rekombinācijas starp alēlēm abos lokusos, gan arī no mutācijām, kas iespējams notikušas katrā no līnijām • r2 ir mazāk atkarīgs no neliela paraugu skaita LD aprēķinos
LD aprēķini – D’ • Divi lokusi, katrā pa divām alēlēm, Aa un B b
LD aprēķini – D’ • D’ vērtība ir starp 0 un 1. D’ = 1 arī tad, ja alēļu frekvences abos lokusos nav pilnīgi identiskas • D’ atkarīgs tikai no rekombinācijas starp alēlēm divos lokusos • D’ nav izmantojams neliela paraugu skaita gadījumā, jo nav iespējams novērot visas 4 iespējamās alēļu kombinācijas. Tas noved pie nepilnīga rekombinācijas novērtējuma un mākslīgi paaugstināta LD
Dabiskās populācijās populāciju struktūru rada ģeogrāfiska norobežotība Lauksaimniecības augos populācijas struktūru nosaka cilvēku selekcijas prakse. Piemēram, vasaras un ziemas šķirnes parasti tiek selekcionētas atsevišķi Abos gadījumos rezultāts ir viens un tas pats – ģenētiska diferenciācija starp populācijām, kas izpaužas kā atšķirīga allēļu frekvenču nostiprināšanās selekcijas rezultātā Populāciju struktūra
Pozitīvas asociācijas var rasties arī populāciju sajaukšanās (admixture) rezultātā Jauktā populācijā jebkura pazīme, kurai ir augstāka frekvence vienā no populācijām būs asociēta ar jebkuru citu allēli, kas arī ir biežāk sastopama šajā populācijā Populāciju struktūras ietekme uz LD
No praktiskā viedokļa, ja ir zināma LD bloku izmērs genomā, ir iespējams paredzēt allēļu genotipu pamatojoties uz citu allēļu genotipu. Piemēram, nosakot dažu lokusu genotipu uz miežu hromosomas 1H, ir iespējams paredzēt miltrasas rezistences specifiskumu LD ir atkarīga no rekombinācijas. Sugās, kur izplatīts inbrīdings, piemēram, pašapputeksnējošos augos, LD bloku izmēri ir lielāki Nelīdzsvarotā saistība kartēšanā
SNP, LD un asociāciju ģenētika Asociāciju kartēšana testē vai kāda SNP’a genotips ir asociēts ar fenotipu. References SNP (sarkani) tiek genotipēti, bet dzelteno SNP genotipu ir iespējams noteikt izmantojot to, ka tie ir LD ar references SNP. Zilo SNP genotipu nav iespējams noteikt, tādēļ to iespējamās fenotipiskās asociācijas nav iespējams noteikt. Kruglyak (2005) Nat Genet 37:1299
Nelīdzsvarotā saistība augu genomā Caldwelletal. (2006) Extreme Population-Dependent Linkage Disequilibrium Detected in an Inbreeding Plant Species, Hordeumvulgare. Genetics, 172: 557
LD bloku izvietojums miežu hromosomās 3H (A) un 2H (B). Augšējā trijstūrī parādīts LD, apakšējā trijstūrī parādītas atbilstošās p vērtības. Marķieru novietojums uz katras hromosomas ģenētiskās kartes parādīts zem A un B paneļa. Krāsu kodējums r2 un p vērtībām parādīts pa labi.
LD bloku izmēri atšķiras dažādās populācijās Mūsdienu miežu šķirnēs (un arī citos lauksaimniecības augos) LD bloku izmēri ir lieli – vairākas cM. Tas saistīts ar selekcijas vēsturi – mūsdienu šķirnes no 20. gs. sākuma šķirnēm šķir nedaudzas paaudzes, tādēļ rekombinācija nav paspējusi samazināt LD bloku izmērus Savvaļas populācijās LD bloku izmēri ir nelieli, jo starp indivīdiem populācijā radniecība ir attāla
LD kartēšanas piemērs – b-glukānu saturs Graudu b-glikānu satura asociāciju kartēšana. b-glukānu saturs noteikts ar Megazyme (MegazymeInternationalIrelandLtd.) izstrādātu enzimātisku metodi Valsts pētījumu programmas agrobiotehnoloģijā ietvaros. SNP marķieri (MAF>0.1) sakārtoti pēc to atrašanās vietas uz miežu ģenētiskās kartes. Asociāciju kartēšana veikta ar programmu TASSEL (Bradburyetal. 2007) izmantojot vispārējo lineāro modeli.
Genomika Fenotips Augu selekcija Ģenētiskā karte Fiziskā karte Genoma secība Alēļu daudzveidība Molekulārie marķieri, genomika un augu selekcija Gēnu funkcija Nils Rostoks
Ģenētiskās kartes modeļorganismos • Ģenētiskās kartes ir noderīgas arī organismos, kuru genoma sekvence ir zināma • Genoma sekvence un informācija par gēniem vēl nav pietiekama, lai pateiktu, kurš gēns nosaka, kuru pazīmi • Ģenētiskai kartēšanaimodeļorganismos ir priekšrocības • - ērti un ātri iespējams radīt lielas kartēšanas populācijas • - pastāv efektīvas molekulāro marķieru sistēmas, kas ļauj veidot detalizētas ģenētiskās kartes • - kad gēns lokalizēt kādā nelielā ģenētiskās kartes intervālā, atliek tikai identificēt tajā esošos gēnus un izvēlēties kandidātgēnus interesējošai pazīmei
Modeļorganismi un lauksaimniecības augi • Lauksaimniecības augi un modeļorganismi ir radniecīgi, piemēram, mieži un kvieši no vienas puses un Brachypodiumdistachyon • Gēni, kas kartēti un klonēti modeļorganismos bieži ir sastopami un veic līdzīgu funkciju arī lauksaimniecības augos • Tādējādi fundamentālās zinātnes pētījumi modeļorganismos ir pielietojami arī lauksaimniecības augu pētījumos un selekcijā
Augu selekcija • Dabiskā izlase – vienas sugas indivīdu atšķirīga spēja izdzīvot un atstāt pēcnācējus • Mākslīgā izlase – cilvēku veikta ar noteiktām īpašībām apveltītu indivīdu izlase • Augu selekcija – ar noteiktām īpašībām apveltītu organismu grupu (šķirņu) mērķtiecīga veidošana • Selekcija un ģenētika ir saistītas. Ģenētika ir selekcijas teorētiskais pamats
Augu šķirnes • InternationUnionfortheProtectionofNewVarietiesofPlants(http://www.upov.int/index_en.html) • Augu šķirne – "a plant grouping within a single botanical taxon of the lowest known rank, which grouping, irrespective of whether the conditions for the grant of a breeder's right are fully met, can be • - defined by the expression of the characteristics resulting from a given genotype or combination of genotypes, • - distinguished from any other plant grouping by the expression of at least one of the said characteristics and • - considered as a unit with regard to its suitability for being propagated unchanged;"
Prasības jaunai augu šķirnei • Formālās prasības • AVS – atšķirība, vienveidība, stabilitāte (DUS – distinctness, uniformity,stability) • SĪN – saimniecisko īpašību novērtēšana • Neformālās prasības • Jaunai šķirnei jābūt apveltītai ar jaunām īpašībām, kādas nepiemīt esošajām šķirnēm, piemēram, augstāka ražība, atšķirīga vai labāka slimību izturība un tmldz.
Prasības jaunai augu šķirnei • Jaunai augu šķirnei jābūt pielāgotai noteiktam mērķim • Piemēram, mieži, kas piemēroti alus gatavošanai satur augstāku cietes un hidrolītisko enzīmu daudzumu, bet zemāku proteīnu daudzumu. Kvieši, kas piemēroti makaronu gatavošanai, neder maizes cepšanai. Miltaini kartupeļi (Lenora, Magdalēna) ir bagāti ar cieti, bet nav piemēroti cepšanai • Augu selekcionāriem tiek izvirzītas prasības veidot šķirnes ar noteiktām īpašībām • Augsta ražība, augsta izturība pret konkrētam reģionam raksturīgām slimībām, ‘laba’ kvalitāte, piemērotība konkrētam tehnoloģiskam procesam
Augu selekcijas metodes • Introdukcija • Izlase (masu izlase un tīro līniju izlase) • Hibridizācija • Inducētās mutācijas • Gēnu inženierija
Dubultotie haploīdi augu selekcijā • Dubultotie haploīdi ir noderīgi ne tikai kartēšanas populāciju veidošanā - lielākā daļa mūsdienu laukaugu šķirņu tiek radītas izmantojot DH tehnoloģiju • DH līnijas ir pilnīgi homozigotiskas, kas paātrina selekcijas procesu un atvieglo šķirņu izmēģinājumus
Gēnu piramidēšana • Vairāku gēnu, kas nosaka līdzīgu fenotipu, apvienošana vienā šķirnē • Piemēram, izturību pret miltrasu (Blumeriagraminisf.sp.hordei) miežos nosaka vairāki gēni. Apvienojot šos gēnus vienā šķirnē iespējams iegūt šķirnes, kas izturīgas pret vairākām miltrasas patogēna rasēm, vai arī tām piemīt kvantitatīvā izturība pret daudzām rasēm • Fenotipiskā atlase gēnu piramidēšanā ir neefektīva, jo nav iespējams atlasīt pēcnācējus ar selekcionāru interesējošām pazīmēm
Divu slimību izturības gēnu piramidēšanas shēma http://www.knowledgebank.irri.org/ricebreedingcourse/Marker_assisted_breeding.htm