Download
1 / 39
390 likes | 550 Views
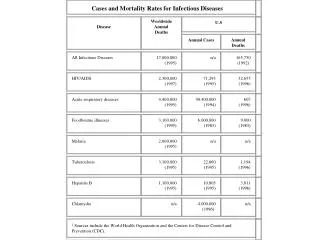
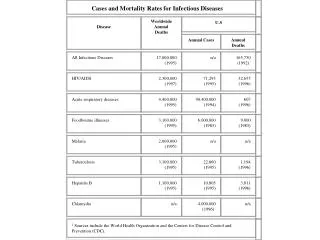
Cases and Mortality Rates for Infectious Diseases. Disease. Worldwide Annual Deaths. U.S. Annual Cases. Annual Deaths. All Infectious Diseases. 17,000,000 (1995). n/a. 165,750 (1992) 2. HIV/AIDS. 2,300,000 (1997). 71,293 (1995). 32,655 (1996). Acute respiratory diseases.

E N D
Cases and Mortality Rates for Infectious Diseases Disease WorldwideAnnualDeaths U.S Annual Cases Annual Deaths All Infectious Diseases 17,000,000(1995) n/a 165,750(1992)2 HIV/AIDS 2,300,000(1997) 71,293(1995) 32,655(1996) Acute respiratory diseases 4,400,000(1995) 90,400,000(1994) 607(1996) Foodbourne illnesses 3,100,000(1995) 6,000,000(1983) 9,000(1983) Malaria 2,000,000(1995) n/a n/a Tuberculosis 3,100,000(1995) 22,860(1995) 1,194(1996) Hepatitis B 1,100,000(1995) 10,805(1995) 3,811(1996) Chlamydia n/a 4,000,000(1996)3 n/a 1 Sources include the World Health Organization and the Centers for Disease Control and Prevention (CDC).
Bacterial Cell Envelopes, LPS, Lipid A partial- structures Immune stimulation- adjuvans- activity Carrier-Targeting Inflammation, Schock Major immunological questions: Endotoxic activities, therapeutic window, routes of administration Routes of activation comparing LPS, uptake by APCs Stimulation of immune responses Possibility for packaging, targeting to specific cells
Detoxified Lipid A monophosphoryl derivative endotoxically inactive acting as an adjuvant inducing tolerance to Salmonella enteritidis LPS and tumor necrosis factor alpha (TNF-alpha) Avanti'sNew Potent Vaccine Adjuvant Natural (Salmonella Minnesota, R595
Limulus Test, Endotoxicity EU/mg S-layer ( B.sphaericus) S-layer ( B. stear.) Ghosts (E.coli O26:B6) Ghosts (S.typhim.) LPS (E.coli O26:B6) LPS ( S. ab. equi ) Control
Endotoxicity does not limit the use of bacterial ghosts as candidate vaccines. Bacterial ghosts prepared from Escherichia coli O26:B6 and Salmonella typhimurium C5 induce dose-dependent antibody responses against bacterial cells or their corresponding lipopolysaccharides (LPS) in doses 25 ng kg-1 when administered intravenously to rabbits. No significant fever responses in rabbits have been recorded in doses of < 250 ng kg-1 E. coli O26:B6 ghosts and up to doses of 250 ng kg-1 S. typhimurium C5 ghosts when applying test methods recommended by the US pharmacopoeia. Vaccine 1997 Feb;15(2):195-202
LPS, CD14 Hypothesis: transmembrane integrin glycoproteins e.g. Mac-1 serve as a signaling partner for glycolipid-linked glycoproteins that lack trans-membrane and cytoplasmic domains.
Inhibition of TNFa by anti CD14 TNF[ng/ml] LPS FCS aCD14 p< (ng/ml) (ug/ml) 10 + - 10 - - 10 + 0,5 0,05 10 - 0,5 10 + 0,1 0,02 10 - 0,1 100 + - 100 - - 100 + 0,5 0,02 100 - 0,5 0,02 100 + 0,1 0,05 100 - 0,1 0,05 - +/- - - +/- +/-
Pathways for LPS stimulation LPS- LBP HSP60? ? IL-6, IFNy,... CD14 GPI Toll- like R-4 CR3 MAC1 ? R Janus Kinase PLC ? Stat 3,1,4 MAPK/ p38 JNK ERK MAPK P CREB, ATF1 NFkB ...,myk,Elk, Mnk eIF4E AH,10/2000
Inhibition of LPS stimulated TNFa Inhibition of Ghost stimulated TNFa by kinase inhibitors in supernatant by kinase inhibitors packed inside ghosts SB/G5 LPS LPS: Co, 0,001; 0,01; 0,1 ug/ml , Co:50-200 pg/ml TNFa, 100%: 5-20 ng/ml TNFa Ghosts: E. Coli, 5, 0,5 ul; 100 %: 5-20 ng/ml TNFa SB203580: 5, 10, 20 uM; G5: Protein kinase CK2 inhibitor, 5,20, 50 uM
Quantitative Real Time PCR( Light Cycler System ) Reverse transcripton reaction RT- PCR: LightCyclerTM System (Roche), FastStart DNA Master SYBR Green I (Roche) Quantification: external cDNA standards with known amounts of initial copy number over a 105 - fold range were coamplified calculation with LightCycler Quantification Software v3.39 during the log- linear phase of the amplification ß- actin was used to normalize for inefficiencies in cDNA synthesis mRNA Isolation: Quick PrepMicro mRNA Purification Kit mRNA semiquantification: Nuleic dotMetricTM Basic Kit (Geno Technology)
Amplification in RT- PCR A B 24 32 30 20 28 26 16 24 22 -dF/dT 20 Fluorescence 12 18 16 106 105 104 103 102 8 14 12 no template 10 4 8 no template 6 0 4 2 0 -4 -2 71 73 75 77 79 81 83 85 87 89 91 93 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 Temperature (°C) Cycle number
IL-12, TNFa mRNA in Ghost- stimulated THP-1 MO (RT- PCR) IL-12 TNFa Eif2-by M Co 3 6 16 16 hrs
Stimulation IL-12 and TNFa in THP1 cellsELISA TNFa LPS IL-12 Ghosts o
Stimulation of IL-12 and TNFa in PBM TNFa IL-12
Immune mediators in DC derived from PBM ( IL4, GMCSF, 5-7 d ) secrete large amounts of inflammatory cytokines as precursors high Ag capture capacity at their immature stage stimulation of quiescient, B and T lymphocytes low levels of antigen to induce strong T cell response
Ghosts and DC What are the mechanisms for the good activation of IL-12 in APC, especially DC ? Does this have a clinical value ?
Microbial stimuli that promote DC maturation Bacteria ( E. coli, M. tuberculosis, Staph. aureus, Strept. ,... Protozoa LPS Bacterial DNA, Viral and doubled-stranded RNA Bacterial heat shock proteins
Activation of DC functions by microbial stimuli. Migration Exit of activated DCs from peripheral sites Entry into the T cell areas of secondary lymphoid tissues Antigen presentation Upregulation of antigen presenting molecules (MHC class I and class II, CD1) Delivery of antigen to the MHC class I pathway Upregulation of molecules involved in interaction with T lymphocytes Costimulatory molecules (B7-1, B7-2) Adhesion molecules (ICAM-1, VLA-4) Signalling molecules (CD40) Production of cytokines Induction of IL-12, TNF, IL-10, IL-6, IFN-/ Recruitment of DC precursors to peripheral sites Increased transient survival of DCs in the absence of growth signals Irreversible maturation followed by apoptotic death
Mature DCs and ghosts induce strong T-cell responses, MLR Pavol KUDELA maturation mix (MM)-TNF-; IL-1; PGE2; IL-6; GM-CSF; IL-4 MM + bacterial ghosts+ GM-CSF + IL-4
Induce ghosts differentiation and activation of DC functions ? MHC class I and II, CD11a,b,c, CD50, CD54, CD58 Activation of T- cells IL-12, T cell attractant chemokine +++ + + +++ + +++
Gut 2000 Jul;47(1):79-87 Non-pathogenic bacteria elicit a differential cytokine response by intestinal epithelial cell/leucocyte co-cultures.Haller D, Bode C, Hammes WP, Pfeifer AM, Schiffrin EJ, Blum S.Institute of Biological Chemistry and Nutrition Science, University Hohenheim, Germany.BACKGROUND AND AIM: Intestinal epithelial cells (IEC) are thought to participate in the mucosal defence against bacteria and in the regulation of mucosal tissue homeostasis Challenge of CaCO-2 cells with non-pathogenic E coli and Lactobacillus sakei induced expression of IL-8, MCP-1, IL-1beta, and TNF-alpha mRNA in the presence of underlying leucocytes. Leucocyte sensitised CaCO-2 cells produced TNF-alpha and IL-1beta whereas IL-10 was exclusively secreted by human peripheral blood mononuclear cells. CaCO-2 cells alone remained hyporesponsive to the bacterial challenge. CONCLUSION: The differential recognition of non-pathogenic bacteria by CaCO-2 cells required the presence of underlying leucocytes. These results strengthen the hypothesis that bacterial signalling at the mucosal surface is dependent on a network of cellular interactions.
Microbiol Immunol 1999;43(10):925-35 Cytokine secretion by stimulated monocytes depends on the growth phase and heat treatment of bacteria: a comparative study between lactic acid bacteria and invasive pathogens.Haller D, Bode C, Hammes WP.Hohenheim University, Stuttgart, Germany. dirk.haller@rdls.nestle.comThe challenge of monocytes with three LAB strains, Listeria monocytogenes or enterohaemorrhagic Escherichia coli (EHEC) elicited a strain specific, dose-dependent biphasic TNF-alpha secretion. LPS exhibited a higher capacity to stimulate monocytes than purified gram positive cell walls or muramyldipeptide. In comparison to pathogenic bacteria, the maximal secretory TNF-alpha response (TNFmax) was up to 2 fold higher with LAB strains. In general, the amount of bacteria (EDmax) necessary to induce maximal TNF-alpha secretion (TNFmax) was approximately 1 to 3 log higher for heat killed bacteria when compared to live bacterial cells illustrating the significant lower potential of heat killed bacteria to activate monocytes.
Bacterial cell envelopes for vaccine development • No endotoxic limitations • lipid A activation pathways, CD14, p38 MAP kinase • provide adjuvans activity comparable to lipid A structures • effective uptake by phagocytes ( others by different mechanisms ?) • potent inducers of IL-12, ( IL-18 ) • may propagate maturation of dendritic cells • may enable packaging of systemic toxic compounds and targeting • to specific cells or organs AH, 10/2000
Ghosts and Immunology Ghosts, preparation and ... M. Szostak Horst Mader M. Schmalnauer Gudrun Kohl Fürst Ladani Beate Mayr Margit Weghofer