Download

1 / 25
250 likes | 386 Views
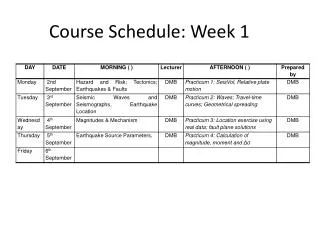
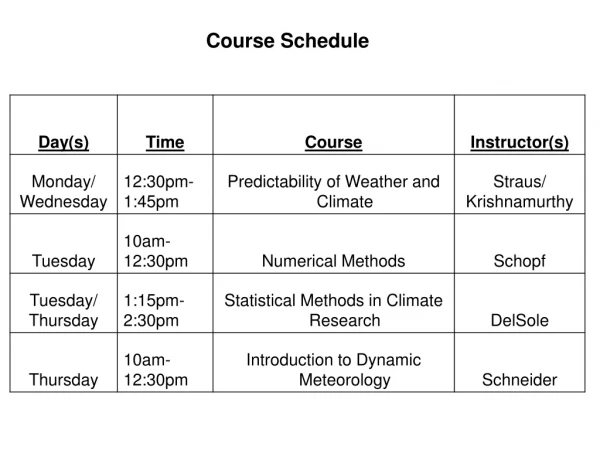
Last week of the course - schedule. Today - lecture Tomorrow- discussion Monday - exam Tuesday - lecture Wednesday - Frank will go over exam (whether graded or not) Thursday - we don’t have the room that day (quarter over) Friday - exam from 3-5 in MS 5200 (sorry about the room).

E N D
Last week of the course - schedule Today - lecture Tomorrow- discussion Monday - exam Tuesday - lecture Wednesday - Frank will go over exam (whether graded or not) Thursday - we don’t have the room that day (quarter over) Friday - exam from 3-5 in MS 5200 (sorry about the room)
The point of the Tps is the discovery that RNA can act as an enzyme with catalytic activity Examples of natural ribozymes Namereactionlocalization Group I self-splicing Nuclear rDNA Tetrahymena Introns organelles, fungi and plants, bacteriophages, bacteria Group II self-splicing organelles, fungi and plants, introns splicing pathway bacteriophages, is identical to bacteria nuclear pre-mRNA introns RNase P 5’ end cleavage Eukaryotes of pre-tRNAs Prokaryotes Ribosome Peptidyl Eukaryotes Transferase Prokaryotes Now discuss how these ribozymes work Uses a co-factor
Group I introns self-splicing Group II introns self-splicing G3’OH A 2’OH G 3’OH A G 3’OH 3’OH Non-lariat intron A 3’OH They only differ in type I uses a free nucleotide as the attacking group 2 mechanisms of self-splicing by ribozymes spliced exon Usual lariat intron
A model of the active Site in a type I reaction Exon1 C G C G C G (The transition state is shown) O U A OH O P O- O internal guide sequence O G C O OH O P O- O O G U Metal binding site O Intron Mg++ O OH G264 O Positions guanosine P O N9 H O N7 N3 OH O H N 2’ H 3’ N N3 N1 O6 1’ H N1 H O 5’ H N9 O H OH O N7 N3 N Guanosine cofactor H N1 C311
Ribosomal RNAs are cleaved out of large precursors in both eukaryotes and bacteria Nucleolar Pol I t-RNAs are made by pol III in eukaryotes and contain introns. (next slide) Bacterial t-RNAs are sometimes cleaved out of precursors
Eukaryotic t-RNA processing (some details later). Probably not co-transcriptional. Note no 5’ cap or 3’ polyA
Both t-RNA and m-RNA go to cytoplasm (already there in bacteria). Now discuss what happens to them - t-RNA first See tp for adapter function of t-RNA
What recognizes the t-RNAs? Use prokaryotic names, mostly. Amino acyl synthetases- they add amino acids and must distinguish different t-RNAs. EF-Tu/Ts- bring all (except initiator) to ribosome so must recognize something common to all. ribosomal binding sites - recognize all and accept delivery. 3 sites in ribosome must be accommodated.
The t-RNAs are chemically modified to allow recognition by diverse molecules
There are non-conserved positions that allow differential recognition by synthetases Added post- transcriptionally There is a constant structure that allows common recognition
Genetic studies indicate that these non-conserved positions are recognized by synthetases
Note that the synthetase does actually bind to these regions Crystal Structure Of E.coli GlnRS- tRNAGln
EF-Tu binds to the opposite surface synthetase
Other factors that must enter the same site are structural mimics The complex fits into the ribosomal A-site.
Charging with amino acids occurs at the added CCA and uses ATP Added post- transcriptionally Amino acids are added here
Aminoacylation (charging) occurs in 2 steps (both require the synthetase) 2-Coupling of the amino acyl adenylate to the 3’OH of the tRNA Aminoacyl-AMP + tRNA Aminoacyl-tRNA + AMP This requires t-RNA but attachment has not occurred. 1-Formation of amino acyl adenylate H O R C C + ATP O- NH3+ O H R C C O Phopho-Ribose-Adenine + PPi NH3+
Another view of aminoacylation of the 3’ terminal adenosine
The fidelity of aminoacylation is checked at each step See next slide 1)binding in the active site is 99% correct 2) After charging, an editing site checks the amino acid “wrongly charged” “rightly charged” does not fit editing site Fits editing site Amino acyl-tRNA Charged tRNA released bond hydrolyzed Uncharged tRNA and amino acid ejected Goes to ribosome Must be re-charged properly
Components of the Ribosome. In addition there are many initiation, elongation and termination factors See Tp and the net reaction Prokaryotes Eukaryotes 50S 60S Large Subunit 2820 kDa 1590 kDa 28S, 5S, 5.8 S rRNAs 23S, 5S rRNAs 49 proteins 31 proteins Peptidyl Transferase Center Peptidyl Transferase Center 30S 40S Small subunit 930 kDa 1400 kDa 16S rRNA 18 S rRNA 21 proteins 33 proteins mRNA Binding proofreading mRNA Binding proofreading
Assembly of an initiation competent translation complex The m-RNA binds to the small subunit (with associated factors). But this is different in bacteria and eukaryotes. In bacteria a guide RNA in the small ribosomal subunit positions the m-RNA. (see next slide)
SD sequences Complementary guide RNA in the 30S subunit 3' end 5' end Translation can start independently at each gene
There is no SD sequence in eukaryotes. Instead the small subunit receives the 5’ cap and scans down to the first initiation codon (AUG). m-RNA joins using 5' cap Scan to AUG
The initiator t-RNA must join to complete the initiation phase. We will only discuss the prokaryotic example (in eukaryotes mostly the names change)
Now this site is ready for the t-RNA that matches the next codon After termination the initiation subunit is freed The initiator t-RNA is brought in after the m-RNA Then the large subunit joins