Download

1 / 38
430 likes | 937 Views
Climate change influence on decomposition and soil microbes. Bob Edmonds. Topics. 1) Evidence for climate change in the Pacific Northwest 2) Influence of climate change on ecosystems 3) Effects of climate change on soil microbes and decomposition 4) Importance of lignicolous fungi

E N D
Climate change influence on decomposition and soil microbes Bob Edmonds
Topics 1) Evidence for climate change in the Pacific Northwest 2) Influence of climate change on ecosystems 3) Effects of climate change on soil microbes and decomposition 4) Importance of lignicolous fungi 5) the diversity of fungi causing wood decay 6) brown versus white rot fungi 7) influence of global warming on wood decay fungi Temperature effects on fungi Influence on wood decay rates 8) Conclusions
1. Evidence for climate change in the west • Temperature and precipitation trends in WA, OR, CA, ID, MT and CO
Precipitation trends – MT, CO MT CO • CO
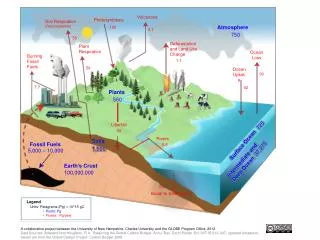
2. Influence of climate change (CO2 and temperature increase and changes in moisture) on ecosystems Ecosystem responses to global warming will be complex and varied (Shaver et al. 2000)
3) Effects of climate change on soil microbes and decomposition
Global change will influence soil ecology: • Directly • - Affecting the growth, survival and dispersal of soil • organisms • - Changing decomposition rates and C and N cycling • Indirectly • - Influencing plant productivity and litterfall rates and • root turnover and soil organic matter • Changing substrate chemistry • Changing plant species composition • Lots of feedback mechanisms
Direct and indirect effects of climate change on soil microbial communities and routes offeedback to global warmingthrough carbon dioxide production. Direct effects include the influence on soil microbes and greenhouse gas production of temperature, changing precipitation and extreme climatic events, whereas indirect effects result from climate-driven changes in plant productivity and vegetation structure which alter soil physicochemical conditions, the supply of carbon to soil and the structure and activity of microbial communities involved in decomposition processes and carbon release from soil Bardgett, Richard D., Freeman, Chris, Ostle, Nicholas J. 2008. ISME JOURNAL 2 (8): 805-814 Microbial contributions to climate change through carbon cycle feedbacks
Elise Pendall, Lindsey Rustad, Josh Schimel. 2008. Towards a predictive understanding of belowground process responses to climate change: have we moved any closer? Functional ecology 22(6): 937-940. Interacting effects of global change forcing factors on belowground processes act directly on plant communities to alter primary productivity, litter quantity and quality, and root production rates. Atmospheric CO2 concentrations have indirect effects on soil moisture, while temperature can alter N and moisture availability. The quantity and quality of soil organic matter and labile root exudates interact with microbial communities to alter rates of biogeochemical cycles. Microbial activity regulates belowground feedbacks to climate change by altering rates of decomposition and reactive N (e.g. N2O) production.
How do you study the effects of global warming? • Soil heating cables or lamps • Closed chambers – CO2, temp, moisture • Open (free air) chambers – CO2 • Lab studies – variable temp, moisture • Mathematical simulation models
Chen, H., Rygiewicz, P. T.Johnson, M. G.Harmon, M. E.Tian, H., Tang, J. W. 2008. Chemistry and long-term decomposition of roots of Douglas-fir grown under elevated atmospheric carbon dioxide and warming conditions JOURNAL OF ENVIRONMENTAL QUALITY 37 (4) : 1327-1336 Elevated atmospheric CO2, concentrations and warming may affect the quality of litters of form plants and their subsequent decomposition in ecosystems, thereby potentially affecting the global carbon cycle. We used small diameter roots of Douglas-fir seedlings that were grown for 4 yr in a 2 x 2 factorial experiment: ambient or elevated (+ 180 ppm) atmospheric CO2 concentrations, and ambient or elevated (+3.8 degrees C) atmospheric temperature. Exposure to elevated CO2 significantly increased water-soluble extractives concentration, but had little effect on the concentration of N, cellulose, and lignin of roots. Elevated temperature increased %WSE and decreased the %lignin content of fine roots. This study indicates that short-term decomposition (< 9 mo) and long-term responses are not similar. It also suggests that increasing atmospheric CO2 had little effect on the carbon storage of Douglas-fir old-growth forests of the Pacific Northwest.
Richard T. Conant, J. Megan Steinweg, Michelle L. Haddix, Eldor A. Paul, Alain F. Plante, Johan Six. 2008. EXPERIMENTAL WARMING SHOWS THAT DECOMPOSITION TEMPERATURE SENSITIVITY INCREASES WITH SOIL ORGANIC MATTER RECALCITRANCE. Ecology: Vol. 89, No. 9, pp. 2384-2391. Soil C decomposition is sensitive to changes in temperature, and even small increases in temperature may prompt large releases of C from soils. Older, more-resistant C fractions may be more temperature sensitive. We incubated soils at constant temperature to deplete them of labile soil OM and then successively assessed the CO2-C efflux in response to warming. We found that the decomposition response to experimental warming early during soil incubation (when more labile C remained) was less than that later when labile C was depleted. These results suggest that the temperature sensitivity of resistant soil OM pools is greater than that for labile soil OM and that global change-driven soil C losses may be greater than previously estimated.
Sokolov, Andrei P., Kicklighter, David W., Melillo, Jerry M., Felzer, Benjamin S., Schlosser, C. Adam, Cronin, Timothy W. 2008. :JOURNAL OF CLIMATE 21(15):3776-3796. The impact of carbon-nitrogen dynamics in terrestrial ecosystems on the interaction between the carbon cycle and climate was studied using an earth system model of intermediate complexity, the MIT Integrated Global Systems Model (IGSM). Simulations show that consideration of carbon-nitrogen interactions not only limits the effect of CO2 fertilization but also changes the sign of the feedback between the climate and terrestrial carbon cycle. In the absence of carbon-nitrogen interactions, surface warming significantly reduces carbon sequestration in both vegetation and soil by increasing respiration and decomposition (a positive feedback). If plant carbon uptake, however, is assumed to be nitrogen limited, an increase in decomposition leads to an increase in nitrogen availability stimulating plant growth. The resulting increase in carbon uptake by vegetation exceeds carbon loss from the soil, leading to enhanced carbon sequestration (a negative feedback). Under very strong surface warming, however, terrestrial ecosystems become a carbon source whether or not carbon-nitrogen interactions are considered. Models that ignore terrestrial carbon-nitrogen dynamics will underestimate reductions in carbon emissions required to achieve atmospheric CO2 stabilization at a given level.
4. Importance of lignicolous fungi As a group, lignicolous fungi play a vital role in recycling the carbon and nutrients locked up in wood. Some species are capable of killing trees. They contribute CO2 to the atmosphere and organic matter to soil, and provide wildlife and plant habitat. They occur in all ecosystems with wood, including deserts Most are saprophytes on dead wood, but many invade living plant to decay heartwood.
5. The diversity of fungi causing wood decay Each has its ecological niche
Taxonomy and numbers of N. American wood-rotting fungi(Gilbertson 1980) Mostly basidiomycetes with a few ascomycetes (Xylariaceae) – may not be able to invade wood without alteration by hyphomycetes, bacteria or yeasts. Basidiomycete orders Tremellales – 101 species Aphyllophorales – 948 species Agaricales – 620 species Total – 1669 species
Others Bird’s nest Puffball
6. Brown versus white rot fungi There are fewer brown rot fungi (106 sp.) than white rot (1563 sp.) – 6% of total They evolved from the white rot fungi. Majority of brown-rot fungi are in the Polyporaceae Most are associated with conifers Initially brown rot fungi cause greater weight loss in wood than white rot fungi.
Brown rot versus white rotDoes it matter?Yes, ecologically http://ocid.nacse.org/classroom/fungi/bot461/
The distribution of North American wood rotting fungi is related to distribution of host plants. However, species of wood decay fungi are generally more widely dispersed than species of higher plants (Gilbertson 1980). But some like E. tinctorium only occur only in the west even though suitable hosts (balsam fir and Fraser fir occur in the east). So far all species are endemic, as far as we know.
7. Influence of global warming on fungi and wood decay rates Effect of temperature on growth and wood decay Effect on fungal fruiting
Temperature effects On average optimum temperature for growth is about 28 C; as low as 20C and has high as 35 C
Temperature accounts for 34% variation in CWD decay rates Mackensen et al. (2003)
Rapid and recent changes in fungal fruiting patterns in Britain Over 56 years – 315 species, 1400 locations First fruiting date is now earlier and last fruiting date is later due to increases in later summer temperature and October rainfall. Many species now fruit in spring as well as fall.
8. Conclusions • We aren’t very good at understanding or predicting belowground process responses to climate change. What will happen to • mycorrhizal fungi? Will their contribution as decomposers • increase? • 2. Global warming will increase decomposition and the decay • rate of woody residues, but changing moisture will also play a role • 3. Fungi will respond quickly to global warming • 4. Fruiting patterns and spore production of wood rotting and other fungal species, including mycorrhizal fungi, will change • 5. Woody residues will continue to decline with • increase use of wood for timber, heating and biofuels • 6. Some species of wood rotting fungi may go extinct, especially • brown rot fungi