
光合作用的过程
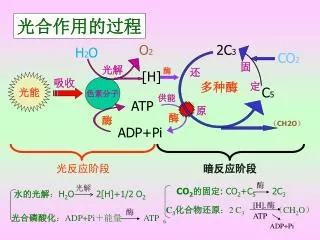
[H],. C 3 化合物还原 : 2 C 3 ( CH 2 O ) 6. 供能. 酶. 光解. 酶. 酶. CO 2 的固定 : CO 2 +C 5 2C 3. 水的光解 : H 2 O 2[H]+1/2 O 2. 光合磷酸化 : ADP+Pi +能量 ATP. ATP. ADP+Pi. 光解. 光合作用的过程. O 2. 2C 3. H 2 O. CO 2. 固 定. 酶. 还 原. [H]. 光能. 吸收. 色素分子. 多种酶.

光合作用的过程
E N D
Presentation Transcript
[H], C3化合物还原:2 C3 (CH2O)6 供能 酶 光解 酶 酶 CO2的固定: CO2+C5 2C3 水的光解:H2O 2[H]+1/2 O2 光合磷酸化:ADP+Pi+能量 ATP ATP ADP+Pi 光解 光合作用的过程 O2 2C3 H2O CO2 固 定 酶 还 原 [H] 光能 吸收 色素分子 多种酶 C5 ATP 酶 酶 (CH2O) ADP+Pi 光反应阶段 暗反应阶段
光合作用的过程和能量转变 • 光合作用的实质是将光能转变成化学能。 根据能量转变的性质,将光合作用分为三个阶段: • 1.原初反应:光能的吸收、传递和转换成电能; • 2.电子传递和光合磷酸化:电能转变为活跃化学能; • 3.碳同化:活跃的化学能转变为稳定的化学能。
表1 光合作用中各种能量转变情况 • 能量转变光能 电能 活跃的化学能 稳定的化学能 • 贮能物质量子电子ATP、NADPH2碳水化合物等 • 转变过程原初反应 电子传递 光合磷酸化 碳同化 • 时间跨度(秒)10-15-10-9 10-10-10-4 100-101 101-102 • 反应部位PSⅠ、PSⅡ颗粒 类囊体膜 类囊体 叶绿体间质 • 是否需光需光不一定,但受光促进不一定,但受光促进
第三节 原初反应 • 原初反应 是指从光合色素分子被光激发,到引起第一个光化学反应为止的过程。 • 它包括: 光物理-光能的吸收、传递 光化学-有电子得失 • 原初反应特点 • 速度非常快,10-12s∽10-9s内完成; • 与温度无关,(77K,液氮温度)(2K,液氦温度); • 量子效率接近1
概念 反应中心色素:少数特殊状态的chl a分子,它具有光化学活性,是光能的“捕捉器”、“转换器”。 聚光色素(天线色素):没有光化学活性,只有收集光能的作用,包括大部分chla 和全部chlb、胡萝卜素、叶黄素。
光合膜上能进行完整光反应的最小结构单位 光合单位
一、光能的吸收与传递 (一) 激发态的形成 • 能量的最低状态─基态。 • 色素分子吸收了一个光子后-----高能的激发态。 • Chl(基态)+hυ 10-15s Chl* (激发态) 图8 叶绿素分子对光的吸收及能量的释放示意图 各能态之间因分子内振动和转动还表现出若干能级。
叶绿素分子受光激发后的能级变化 • 叶绿素: 红光区:被红光激发,电子跃迁到能量较低的第一单线态 蓝光区:被蓝光激发,电子跃迁到第二单线态。 • 配对电子的自旋方向:单线态;三线态;第一单线态;第二单线态 图8 叶绿素分子对光的吸收及能量的释放示意图虚线表示吸收光子后所产生的电子跃迁或发光, 实线表示能量的释放, 半箭头表示电子自旋方向
(二)激发态的命运 1.放热 2.发射荧光与磷光 3.色素分子间的能量传递 4.光化学反应
1.放热 • 激发态的叶绿素分子在能级降低时以热的形式释放能量,此过程又称内转换或无辐射退激。 Chl* →Chl+热 Chl* → ChlT +热 ChlT → Chl+热 • 另外吸收蓝光处于第二单线态的叶绿素分子,其具有的能量虽远大于第一单线态的叶绿素分子。但超过部分对光合作用是无用的,在极短的时间内以热能释放。 • 由于叶绿素是以第一单线态参加光合作用的。在能量利用上蓝光没有红光高。 ??
2. 发射荧光与磷光 • 激发态的叶绿素分子回至基态时,可以光子形式释放能量。 • 荧光。 • 磷光。 • Chl* Chl + hν 荧光发射 • ChlT Chl + hν 磷光发射 • 磷光波长比荧光波长长,转换的时间也较长,而强度只有荧光的1%,故需用仪器才能测量到。 10-9s 10-2s
对提取的叶绿体色素浓溶液照光,在与入射光垂直的方向上可观察到呈暗红色的荧光。对提取的叶绿体色素浓溶液照光,在与入射光垂直的方向上可观察到呈暗红色的荧光。 离体色素溶液为什么易发荧光? • 因为溶液中缺少能量受体或电子受体的缘故。 • 荧光猝灭剂:在色素溶液中,如加入某种受体分子,能使荧光消失。常用Q表示。在光合作用的光反应中,Q即为电子受体。 • 色素发射荧光的能量与用于光合作用的能量是相互竞争的,这就是叶绿素荧光常常被认作光合作用无效指标的依据。
3.色素分子间的能量传递 • 激发态的色素分子把激发能传递给处于基态的同种或异种分子而返回基态的过程。 Chl*1+ Chl2 Chl1+Chl*2 供体分子 受体分子 • 一般认为,色素分子间激发能不是靠分子间的碰撞传递的,也不是靠分子间电荷转移传递的,可能是通过“激子传递”或“共振传递”方式传递。
激子传递 • 激子通常是指非金属晶体中由电子激发的量子,它能转移能量但不能转移电荷。 • 这种在相同分子内依靠激子传递来转移能量的方式称为激子传递。
共振传递 • 在色素系统中,一个色素分子吸收光能被激发后,其中高能电子的振动会引起附近另一个分子中某个电子的振动(共振),当第二个分子电子振动被诱导起来,就发生了电子激发能量的传递。这种依靠电子振动在分子间传递能量的方式就称为“共振传递”。 共振传递示意图 • 在共振传递过程中,供体和受体分子可以是同种,也可以是异种分子。分子既无光的发射也无光的吸收,也无分子间的电子传递 。
通过上述色素分子间的能量传递,聚光色素吸收的光能会很快到达并激发反应中心色素分子,启动光化学反应。通过上述色素分子间的能量传递,聚光色素吸收的光能会很快到达并激发反应中心色素分子,启动光化学反应。 图 光合作用过程中能量运转的基本概念
二、光化学反应 (一)反应中心与光化学反应 1.反应中心 • 原初反应的光化学反应是在光系统的反应中心进行的。 • 反应中心是发生原初反应的最小单位。 • 由反应中心色素分子、原初电子受体、次级电子受体与供体等电子传递体,以及维持这些电子传递体的微环境所必需的蛋白质等成分组成的。
反应中心中的原初电子受体是指直接接收反应中心色素分子传来电子的电子传递体 • 反应中心色素分子是光化学反应中最先向原初电子受体供给电子的,因此反应中心色素分子又称原初电子供体。
去镁叶绿素 去镁叶绿素 副叶绿素 副叶绿素 胡萝卜素 配对叶绿素 光系统‖的反应中心
2.光化学反应 • 原初反应的光化学反应实际就是由光引起的反应中心色素分子与原初电子受体间的氧化还原反应,可用下式表示光化学反应过程: P·A hυ P*·A P+·A- 基态反应中心 激发态反应中心 电荷分离的反应中心 • 反应中心出现了电荷分离(P+)(A-) ,到这里原初反应也就完成了。
原初电子供体P+失去电子,有了“空穴”,成为“陷阱” ,便可从次级电子供体(D)那里争夺电子;而原初电子受体得到电子,使电位值升高,供电子的能力增强,可将电子传给次级电子受体(A1) 。 • 那么电荷分离后反应中心的更新反应式可写为: • D·〔P+·A-〕·A1 D+·〔P·A〕·A1- • 这一过程在光合作用中不断反复地进行,从而推动电子在电子传递体中传递。
(二)PSⅠ和PSⅡ的光化学反应 • 高等植物的两个光系统有各自的反应中心。 • PSⅠ和PSⅡ • P700、P680 红降现象和双光增益效应
PSⅠ和PSⅡ反应中心中的原初电子供体很相似,都是由两个叶绿素a分子组成的二聚体,分别用P700、P680来表示。 • 这里P代表色素,700、680则代表P氧化时其吸收光谱中变化最大的波长位置是近700nm或680nm处。 图9 菠菜反应中心色素吸收光谱的变化 照光下PSⅠ(A)、PSⅡ(B)反应中心色素氧化,其氧化态,与还原态的吸收光谱差值最大变化的波长所在位置分别是700nm(A)和682nm(B)。
光合作用的两个光系统和电子传递方案 吸收红光的光系统Ⅱ(PSⅡ)产生强氧化剂和弱还原剂。吸收远红光的光系统Ι(PSΙ)产生弱氧化剂和强还原剂。PSⅡ产生的强氧化剂氧化水,同时,PSΙ产生的强还原剂还原NADP+。
PSⅠ和PSⅡ的光化学反应 • PSⅠ的原初反应: • P700·A0hυ P700*·A0 P700+·A0- • PSⅡ的原初反应: • P680·Pheo hυ P680*·Pheo P680+·Pheo- • 在原初反应中,受光激发的反应中心色素分子发射出高能电子,完成了光→电转变,随后高能电子将沿着光合电子传递链进一步传递。
第四节 电子传递和光合磷酸化 原初反应的结果: • 产生了高能电子推动着光合膜上的电子传递。 电子传递的结果: • 引起水的裂解放氧以及NADP+的还原; • 建立了跨膜的质子动力势,启动了光合磷酸化,形成ATP。 这样就把电能转化为活跃的化学能。
一、电子和质子的传递 (一)光合链 • 指定位在光合膜上的,由多个电子传递体组成的电子传递的总轨道。 • 希尔(1960)等人提出并经后人修正与补充的“Z”方案。 • 即电子传递是在两个光系统串联配合下完成的,电子传递体按氧化还原电位高低排列,使电子传递链呈侧写的“Z”形。
“Z”方案特点: • (1)电子传递链主要由光合膜上的PSⅡ、Cyt b6/f、PSⅠ三个复合体串联组成。 • (2)电子传递有二处逆电势梯度,即P680至P680*,P700至P700*,逆电势梯度的电子传递均由聚光色素复合体吸收光能后推动,而其余电子传递都是顺电势梯度的。 图10 叶绿体中的电子传递模式 方框代表了蛋白复合物。LHCⅠ和LHCⅡ分别是PSⅠ和PSⅡ各自的聚光色素复合体,M为含Mn的放氧复合体,实线箭头表示非环式电子传递方向;虚线箭头表示环式或假环式电子传递分叉处。
(3)水的氧化与PSⅡ电子传递有关,NADP+的还原与PSⅠ电子传递有关。电子最终供体为水,水氧化时,向PSⅡ传交4个电子,使2H2O产生1个O2和4个H+。电子的最终受体为NADP+。 • (4)PQ是双电子双H+传递体,它伴随电子传递,把H+从类囊体膜外带至膜内,连同水分解产生的H+一起建立类囊体内外的H+电化学势差。
(二)光合电子传递体的组成与功能 1.PSⅡ复合体 • PSⅡ的生理功能是吸收光能,进行光化学反应,产生强的氧化剂,使水裂解释放氧气,并把水中的电子传至质体醌。 (1)PSⅡ复合体的组成与反应中心中的电子传递 • PSⅡ是含有多亚基的蛋白复合体。组成:聚光色素复合体Ⅱ、中心天线、反应中心、放氧复合体、细胞色素和多种辅助因子。 PSII反应中心结构模式图
组成中心天线的CP47和CP43是指分子量分别为47 000、43 000并与叶绿素结合的聚光色素蛋白复合体,它们围绕P680,比LHCⅡ更快地把吸收的光能传至PSⅡ反应中心,所以被称为中心天线或“近侧天线”。
PSⅡ反应中心的核心部分是分子量分别为32 000和34 000的D1和D2两条多肽。D1很容易受到光化学破坏,会发生活性逆转。 反应中心的次级电子供体Z、中心色素P680、原初电子受体Pheo、次级电子受体QA、QB等都结合在D1和D2上。 其中与D1结合的质体醌定名为QB,与D2结合的质体醌定名为QA。Q有双重涵义,既是醌(quinone)的字首,又是荧光猝灭剂(quencher)的字首。
QA是单电子体传递体,每次反应只接受一个电子生成半醌。它的电子再传递至QB,QB是双电子传递体,QB可两次从QA接受电子以及从周围介质中接受2个H+而还原成氢醌(QH2 )。这样生成的氢醌可以与醌库的PQ交换,生成PQH2。
(2)水的氧化与放氧 • 放氧复合体(OEC)又称锰聚合体(M,MSP),在PSⅡ靠近类囊体腔的一侧,参与水的裂解和氧的释放。 • 水的氧化反应是生物界中植物光合作用特有的反应,也是光合作用中最重要的反应之一。 • 每释放1个O2需要从2个H2O中移去 4 个 e-,同时形成 4 个 H+。 • CO2+2H2O*光 叶绿体(CH2O)+ O2*+ H2O
20世纪60年代,法国的乔利尔特(P. Joliot)发明了能灵敏测定微量氧变化的极谱电极,用它测定小球藻的光合放氧反应。 • 他们将小球藻预先保持在暗中,然后给以一系列的瞬间闪光照射(如每次闪光5~10μs,间隔300ms)。 • 发现闪光后氧的产量是不均量的,是以4为周期呈现振荡,即第一次闪光后没有O2的释放,第二次释放少量O2,第三次O2的释放达到高峰,每4次闪光出现1次放氧峰 • 用高等植物叶绿体实验得到同样的结果。 图12 系列闪光对小球藻放氧量的影响 在第三个闪光阶段氧形成量最大,以后每四个闪光都可以看到一个周期性的峰值。氧形成量大约在第20个闪光后体系放O2的周期性会逐渐消失,放O2量达到某一平稳的数值。(Joliot,1965 )
科克(B.Kok,1970)等人根据这一事实提出了关于H2O裂解放氧的“四量子机理假说”: • ①PSⅡ的反应中心与H2O之间存在一个正电荷的贮存处(S) • ②每次闪光,S交给PSⅡ反应中心1个e-; • ③当S失去4e-带有4个正电荷时能裂解2个H2O释放1个O2(图13), • 图中S即为M,按照氧化程度(即带正电荷的多少)从低到高的顺序,将不同状态的M分别称为S0、S1、S2、S3和S4。即S0不带电荷,S1带 1 个正电荷,……S4带4个正电荷。 • 每一次闪光将状态 S 向前推进一步,直至S4。然后S4从 2 个 H2O中获取 4 个e-,并回到S0。 图13 在水裂解放氧中的S状态变化 (修改过的Kok水氧化钟模型,注明了H+的释放部位)
质醌(PQ)也叫质体醌,是PSⅡ反应中心的末端电子受体,也是介于PSⅡ复合体与Cyt b6/f复合体间的电子传递体。 • “PQ库” ; 叶绿素的5%~10%。 • 质体醌为脂溶性分子,质体醌有一个醌的头和一个长的非极性的尾,尾部使质体醌定位于膜中,能在类囊体膜中自由移动,转运电子与质子。 2.质醌
PQ库作为电子、质子的缓冲库,能均衡两个光系统间的电子传递(如当一个光系统受损时,使另一光系统的电子传递仍能进行),可使多个PSⅡ复合体与多个Cyt b6/f 复合体发生联系,使得类囊体膜上的电子传递成网络式地进行。 • 质体醌是双电子、双质子传递体,对类囊体膜内外建立质子梯度起着重要的作用。 Cytb6/f