Download

1 / 1
10 likes | 190 Views
Fig. 7. Fig. 9. Fig. 6. Fig. 8. Fig. 5. Fig.1. Fig. 3. Fig. 2. Fig. 4. THE STRUCTURAL CHARACTERISATION OF TWO HOLLIDAY JUNCTIONS Benjamin C. Gale , James H. Thorpe, Susana C. M. Teixeira and Christine J. Cardin The University of Reading Chemistry Department. Introduction

E N D
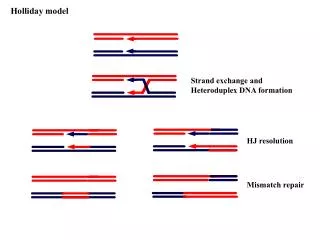
Fig. 7 Fig. 9 Fig. 6 Fig. 8 Fig. 5 Fig.1 Fig. 3 Fig. 2 Fig. 4 THE STRUCTURAL CHARACTERISATION OF TWO HOLLIDAY JUNCTIONS Benjamin C. Gale , James H. Thorpe, Susana C. M. Teixeira and Christine J. Cardin The University of Reading Chemistry Department Introduction The four-way junction, or the Holliday junction plays an important role in the physiology of cells, acting as a central recombination intermediate for many recombination enzymes1. These enzymes promote or catalyse branch migration allowing the exchange of genetic information between DNA duplexes. The Holliday junction is formed through strand exchange of two homologous DNA molecules to give a four-way junction at the cross-over2. As such the Holliday junction represents a structurally distinctive motif that must be recognised at the molecular level by the enzymes through a currently unidentified mechanism. Here we present the single crystal structures for two DNA Holliday junctions formed by d(CCGGTACCGG)23 [1]and d(TCGGTACCGA)2 [2] at 2.35Å and 1.8Å resolutions respectively in the monoclinic space group C2 and discuss the essential features in the stabilisation of the Holliday junction. Data Collection and Processing d(CCGGTACCGG) Temperature: 100K Cryobuffer: Perfluoropolyether (RS 3000) oil Instrument: MAR345 image plate X-ray Source: Synchrotron radiation 1.073Å, X31 beamline DESY, Hamburg Germany Space group: C2 Cell: a = 64.9Å b = 25.4Å c = 37.4Å = 110.6o No. of Observed Reflections: 4466 No. of Unique Reflections: 2390 R-factor: 22.64 Rmerge: 0.056 (0.230) Completeness: 96.41 Average B-factor: 39.94 Å2 Resolution Limits: 30.429 - 2.350 Software: DENZO and SCALEPACK Data Collection and Processing d(TCGGTACCGA) 100K :Temperature Perfluoropolyether (RS 3000) oil :Cryobuffer CCD detector MAR-Research :Instrument Synchrotron radiation 0.8068Å, X11 beamline :X-ray Source DESY, Hamburg Germany C2 :Space group a = 66.1Å b = 23.8Å c = 73.8Å = 110.58 :Cell 9220 :No. of Observed Reflections 8780 :No. of Unique Reflections 22.82 :R-factor 0.0703 (0.2025) :Rmerge 97.35 :Completeness 23.5Å2 :Average B-factor 69.007 - 1.850 :Resolution Limits MOSFLM and CCP4 :Software Crystallisation Conditions d(CCGGTACCGG)2 d(TCGGTACCGA)2 Temperature: 290K 290K Crystallising Solution: 40mM sodium cacodylate (pH 7.0) 75mM sodium cacodylate (pH 7.0) 12mM spermine 35mM calcium chloride 80mM potassium chloride 2.5% MPD 10% MPD Equilibrated against 30% MPD Equilibrated against 35% MPD DNA: 1mM 1mM Method: Sitting Drop Vapour Diffusion Sitting Drop Vapour Diffusion Table 1 A summary of related native decamer sequences In this work the asymmetric unit is either a full (Fig. 1) or half stacked-X (Fig. 2) with a two fold symmetry at its centre. In the case of the full quadruplex the c-axial length is halved and the approximate two-fold symmetry is lost. In either case however, the packing is remarkably similar with helices crossing at approximately 40o in each case (Table 1). The strand exchange at the centre of the structures leads four phosphates to be closely packed (Figs. 3 & 4), thus producing a region of highly negative electrostatic potential. Here the better resolution, when compared with [1] and other Holliday sequences in Table 1 allows two partially hydrated sodium ions near to the phosphate junction in the minor and major groove (Fig. 5) to be defined. Four distorted pentagonal bipyramidal calcium atoms (Fig. 6) anchored in the minor grooves to the G/C base pairs prior to the terminal A/T sites can also be observed. It is apparent that these sites aid in stabilising the solvent structure throughout the minor groove. In addition to these stabilisation forces, well determined solvent bridging sites between the phosphate oxygens at the cross-over on the major groove face and the purine bases either side are present in [2] (Fig 7). For both structures, the minor groove face at the cross-over shows direct interactions between the cytosine N4 sites and the oxygens of the closely packed phosphates (Figs. 8 & 9). Previously4 the ACC step has been shown vital for the formation of the Holliday junction within a crystalline lattice. This again has been demonstrated but further to this we have shown the presence of a purine base either side of the cytosines to be essential in the water mediated stabilisation at the cross-over. Finally the ellusive ion structure of the Holliday sequence has been determined with a clarity highlighting the importance of these sites in the stabilisation of the solvent structure throughout the Holliday structure. References 1. D. M. Lilley & M. F. White (2001). Nature Mol. Cell Biol. 2, 433-44 2. R. Holliday (1964). Genet. Res. 5, 283-304 3. J. H. Thorpe, S. C. M. Teixeira, B. C. Gale & C. J. Cardin (2002). Acta Cryst. D58, 567-569. 4. B. F. Eichman, B. H. M. Mooers, J. M. Vargason & P. Shing-Ho (2000). Proc. Natl Acad. Sci. USA. 97, 3971-3976 5. M. Oritz-Lombardia, A. Gonzalez, R. Eritja, J. Aymani, F. Azorin & M. Coll (1999). Nature Struct Biol. 6, 913-917 6. A. A. Wood, C. M. Nunn, O. J. Trent & S. Niedle (1997). J. Mol. Biol. 269, 827-841 Acknowledgements BCG would like to thank the Association of International Cancer Research for their essential funding and to JHT, SCMT and JHT for their guidance. SCMT is grateful to the Chemistry Department of the University of Reading and the Portuguese Foundation for Science and Technology