Download
1 / 1
10 likes | 121 Views
Investigating the impact of thyroid hormone levels on transcriptional responses in axolotl metamorphosis through microarray analysis. Salamanders serve as cadmium accumulators, making them ideal for biomonitoring. The study uses genomic approaches to understand the molecular mechanisms underlying metamorphosis.

E N D
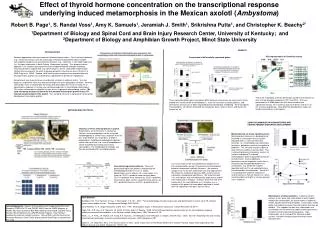
Effect of thyroid hormone concentration on the transcriptional response underlying induced metamorphosis in the Mexican axolotl (Ambystoma) INTRODUCTION Genomic approaches offer great potential to biomonitoring studies. First, functional genomics (e.g., microarray analysis) can be used to gain mechanistic perspective about complex transcriptional responses to environmental toxicants (e.g., cadmium) in non-model organisms (i.e., the tiger salamander in North Dakota, Ambystoma tigrinum). This discovery-driven approach is an excellent tool especially when complemented with complex laboratory functional genomics experiments (e.g. qPCR analysis) on the salamander model axolotl (Ambystoma mexicanum) for which a growing genomics infrastructure exists (Smith et al., 2005; Page et al., 2008). Second, while most genomics projects have provided primarily descriptive data, projects can use genomics approaches to directly test hypotheses. Environments can show continuous and discrete variation in cadmium levels. Very few organisms experience more than one level of exposure to this geographic variation. However, salamanders have a metamorphic life history with two stages that results in exposure to aquatic-borne cadmium (in larvae) and soil-based cadmium (in transformed salamanders). This makes salamander an excellent system for use in genomic biomonitoring studies. We used microarray analysis to determine the standard transcription profile across TH-induced metamorphosis in the axolotl. This standard will serve in comparison to disrupted-development transcription profiles. Generalized regression patterns. These are generalized regression patterns recovered by the methodology describe in Liu et al. (2005). Abbreviations are as follows: LD = linear down, LU = linear up, QLCD = quadratic linear concave down, QLCU = quadratic linear concave up, QLVD = quadratic linear convex down, QLVU = quadratic linear convex up, QC = quadratic concave, and QV = quadratic convex. Salamanders as bioaccululators. Cadmium content analysis of soil, water and salamanders in each habitat indicate that salamanders are accumulators of cadmium. In both aquatic and terrestrial habitats, salamanders exhibit higher liver cadmium content than the habitats that they are found in. This supports the idea that salamanders are accumulators of environmental cadmium and represent an excellent and novel biomonitoring species. Tiger salamanders are at the top of the food web in both systems. No other vertebrate species exhibits these characteristics. Influence of TH on metamorphosis in axolotl. Salamanders are immersed in T4 (thyroxine) solution, and metamorphosis can be assessed morphologically in several ways: reduction of tail fin, loss of gill fimbrae, loss of gill bars, change in pigmentation. Additional morphological indicators can be later assessed, e.g., cranial morphogenesis can be inspected by clearing and staining salamanders. The morphogenetic changes can then be correlated with transcription profiles. Metamorphosis as stress signalling event. Metamorphosis represents a developmental threshold in salamanders that is thyroid-regulated and is a stressful life-history transition, i.e., metamorphosing salamanders lose mass, dehydrate, and are susceptible to predation. This transition may be subject to more intensive cadmium accumulation. This is interesting because microarray analysis of axolotls undergoing TH-induced metamorphosis down-regulate MT during the transformation. Several additional biomarkers may be especially useful then in characterizing disrupted development in salamanders, e.g, keratin 6A is down-regulated and hsp70 is up-regulated across the metamorphic transformation. Keratin 6A is associated with cadmium-induction of several cancers in human (Somji et al., 2004) and transcription of hsp70 is a known general stress response. Robert B. Page1, S. Randal Voss1, Amy K. Samuels1, Jeramiah J. Smith1, Srikrishna Putta1, and Christopher K. Beachy2* 1Department of Biology and Spinal Cord and Brain Injury Research Center, University of Kentucky; and 2Department of Biology and Amphibian Growth Project, Minot State University RESULTS PCA representation of GeneChip results. Comparison of differentially expressed genes. These are scatterplots of the 30 GeneChips based on the rotations of their first two principal components (PC1 and PC2). PCA was performed on all 3688 probe-sets that were available from significance testing. PC1 and PC2 account for 89.2% and 6.2% of the variance respectively. Cano-Martinez development stages are listed in parentheses in the legend. These recovered gene expression profiles differ because some genes are expressed in larval amphibians and turned off at metamorphosis, some are turned on at metamorphosis, and some genes are only up- or down-regulated during metamorphic remodeling. For the purposes of presentation, LD, QCLD, and QLVD are lumped as “down” and LU, QLVU, and QLCU are lumped as “up.” METHODS AND PROTOCOL USING SALAMANDER IN BIOMONITORING AND CHARACTERIZING DISRUPTED DEVELOPMENT Our results allow for the first bioinformatic comparison of transcription profiles during TH-induction of metamorphosis between salamander and frog (Xenopus). We chose to compare our results with salamander skin and frog intestine. We chose this comparison because both systems feature substantial remodeling of ephithelium. We found that only 13% of the genes that were differentially expressed in axolotl were expressed in Xenopus. Of these, fewer than half were expressed in the same direction as in Xenopus, e.g., a majority of the genes that were down-regulated in axolotl were up-regulated in Xenopus and vice versa. REFERENCES Buchholz, D.R., R.A. Heimeier, B. Das, T. Washington, Y.-B. Shi. 2007. Pairing morphology with gene expression and identification of a core set of TH-induced genes across tadpole tissues. Developmental Biology 303:576-590. Cano-Martinez, A., A. Vargas-Gonzalez, and M. Asai. 1994. Metamorphic stages in Ambystoma mexicanum. Axolotl Newsletter 23:64-71. Page, R.B., S.R. Voss, A.K. Samuels, J.J. Smith, S. Putta, and C.K. Beachy. 2008. Effect of thyroid hormone concentration on the transcriptional response underlying induced metamorphosis in the Mexican axolotl (Ambystoma). BMC Genomics 9:78. Smith, J.J., S. Putta, J.A. Walker, D.K. Kump, A.K. Samuels, J.R. Monaghan, D.W. Weisrock, C. Staben, and S.R. Voss. 2005. Sal-site: integrating new and existing ambystomatid salamander research and informational resources. BMC Genomics 6:181. Somji, S., J.H. Todd, M.A. Sens, S.H. Garrett, and D. A. Sens. 2000. Expression of Heat Shock Protein 60 in Human Proximal Tubule Cells Exposed to Heat, Sodium Arsenite and CdCl2. Toxicol Lett. 115:127-136. Acknowledgments. Aspects of this projects were supported by NIH Grant Numbers P20RR-016741 and P20RR-16481 from the INBRE program of NCRR, NIH Grant Number 5-R24-RR016344-06 from NCRR, and NSF Grant Number IBN-0242833 of the CAREER Award Program. Ryan Winburn provided the cadmium content analyses. Donna Walls and the University of Kentucky Microarray Core Facility processed the GeneChips associated with this study.