Download
1 / 37
370 likes | 597 Views
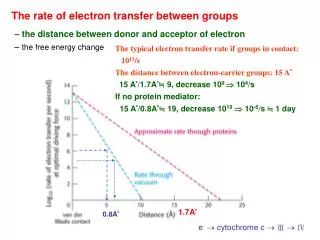
The rate of electron transfer between groups. The typical electron transfer rate if groups in contact: 10 13 /s The distance between electron-carrier groups: 15 A ° 15 A°/1.7A ° ≒ 9, decrease 10 9 10 4 /s If no protein mediator: 15 A°/0.8A°≒ 19, decrease 10 19 10 -6 /s ≒ 1 day.

E N D
The rate of electron transfer between groups The typical electron transfer rate if groups in contact: 1013/s The distance between electron-carrier groups: 15 A° 15 A°/1.7A°≒ 9, decrease 109 104/s If no protein mediator: 15 A°/0.8A°≒ 19, decrease 1019 10-6/s ≒ 1 day – the distance between donor and acceptor of electron – the free energy change 1.7A° 0.8A° e- cytochrome c Ⅲ Ⅳ
The rate of electron transfer between groups – the distance between donor and acceptor of electron – the free energy change inverted region Ch. 19 ?
Chemiosmotic hypothesis (1961, Mitchell P.) NADH oxidation ADP phosphorylation a covalent high-energy intermediate or an activated protein conformation proton-motive force drives the synthesis of ATP bymitochondrial ATP synthase(F1F0ATPase, complex Ⅴ) Proton-motive force ﹙ p﹚: the pH gradient ﹙ pH﹚ + the charge gradient [membrane potential ﹙﹚]
Testing the chemiosmotic hypothesis Artificial membrane A separate system: (A purple-membrane protein, pump protons when light) Respiratory chain Proton gradient ATP synthase (from beef heart)
19.2 ATP synthesis Mitchell: chemiosmotic model, proton-motive-force ADP + Pi + n Hp+ → ATP + H2O + n HN+ Nelson a proton pore associated with ATP synthase
ATP synthase mechanism: Mg2+ require Orthophosphate HPO42-
ADP + Pi + ATP synthase in H218O isotopic-exchange experiments: enzyme-bound ATP forms readily in the absence of a proton-motive force + ATP synthase The role of proton gradient is to release ATP from ATP synthase but not to form ATP
ATP synthase structure proton channel complex c-ring (10~14 c subunits) 1a, 2b, 1 matrix (3, 3, , and ) subunits participate directly in ATP synthesis Two functional components: 1. Moving units c-ring, stalk 2. Stationary unit
Boyer PD (2000): binding-change mechanism subunit make the 3 subunits unequivalent Open The conformation of releasing ATP Loss The conformation of binding ADP and Pi Tight The conformation of binding ATP
Binding-change mechanism: A counterclockwise direction in subunit T L Proton drive O
The smallest molecular motor Only cloned 33 subunits Fluorescence labeled Polyhistidine tags of N-terminal of subunit Nicklel ions are coated on glass surface The subunit was rotated, driven by the hydrolysis of ATP 120° increment/ATP hydrolysis Near 100% efficiency
Components of the proton-conducting unit of ATP synthase The site of proton entrance Asp61 COO-/COOH a pair of -helices Two hydrophilic half-channel Do not span the membrane Directly about one c subunit, separately
Proton flow/c-ring rotation power rotation, then ATP synthesis [H+]cyto/[H+]matrix 25 hydrophobic interaction Arg210 in subunit a(02), Ex. 18
Proton path through the membrane C ring tightly links to and subunits C ring rotate rotate 360°/3 ATP 10 c subunits/3 ATP 3.33 protons/ATP H+ NADH: 10 H+, 2.5 ATP FADH2: 6 H+, 1.5 ATP Ex. 19
116 watts (joule/s) provides energy to sustain a resting person 921 earthquake
§ 18.5 Shuttles: an array of membrane-spanning transporter proteins Glycerol 3-phosphate shuttle: electrons of cytosolic NADH from glycolysis enter mitochondrial electron transport chain especially prominent inmuscle and some insects lack lactate dehydrogenase G3P 1,3bisP against NADH gradient 1.5 ATP formation for 1 NADH from glycolysis
Malate-aspartate shuttle in heart and liver is readily reversible the NADH/NAD+ ratio of the cytosol is higher than that of mitochondria 2.5 ATP formation for 1 NADH from glycolysis transamination transamination
ATP-ADP translocase(adenine nucleotide translocase or ANT) highly abundant in the inner mitochondrial membrane (15%) 30 kd, a single nucleotide-binding site, without Mg2+﹙Ex. 20﹚ ADP first entry then coupled to ATP exit, even though the transport rate of ATP is 30-fold higher than that of ADP high energy consumed, about ¼ of the energy from e- transfer atractyloside P site N site Membrane potential Proton-motive force Ex. 22 Bongkrekic acid
Mitochondrial transporters H2PO4- ATP synthasome: ATP synthase, ATP-ADP translocase, phosphate carrier (electroneutral exchange) Dicarboxylate carrier 40 genes in human genome are encoded
Consumed a proton during ATP translocation Other metabolites translocated 2e- transferless than 10H+ formation Glucose is completely oxidized Glycolysis + 2 TCA cycle (GTP) + 2 or +5 or 32 anaerobic metabolism: 2 ATP
Respiratory control (or acceptor control) the rate of oxidative phosphorylation is primarily regulated byADP level the rate of oxidative phosphorylation is determined bythe need for ATP
Energy charge regulation The rate of TCA cycle is controlled by the availability of NAD+ and FAD ATP synthasome
Uncoupling proteins (UCPs) dissipate proton flow generate heat UCP1 (thermogenin) Temp. -adrenergic agonists triacylglycerol degrade free fatty acids liberate Activate UCP-1
UCP proteins: generate heat to maintain body temperature in hibernating animals, some newborn animals, and in mammals adapted to cold Brown adipose tissue (also brown fat mitochondria), which is very rich in mitochondria, is specialized for nonshivering thermogenesis (vs. white adipose tissue) regulate the body weight (obesity)[UCP2 and UCP3] increase the evaporation of odoriferous molecules, skunk cabbage p. 533 7th line: greenish-colored cytochromes
Uncoupler disrupted the coupling of electron transport and phosphorylation – dissipated proton-motive force oxygen consumption, NADH oxidation, no ATP formation heat loss • DNP and certain other acidic aromatic compounds used in some herbicides, fungicides, weight- loss drug (?)
Sites of action of inhibitors of electron transport ferric ferrous form of heme a3
Oligomycin Dicyclohexylcarbodiimide (DCCD) prevent the influx of protons through ATP synthase
Nelson Alternative mechanisms in plant mitochondria Araceae: one family of stinking plants thermogenesis a cyanide-resistant QH2 oxidase bypass complex III and Ⅳ a rotenone insensitive NADH dehydrogenase, bypass complex Ⅰ a skunk cabbage
Mitochondria ¤ semiautonomous organelles ¤ endosymbiotic double membrane, circular DNA, specific transcription and translation machinery ¤ Maternally inherited
Mitochondria – diseases a center of energy metabolism Leber hereditary optic neuropathy – NADH-Q oxidoreductase (complexⅠ) mutation – resulted in blindness during midlife chance fluctuation percentage the threshold of defect the accumulation of mutations effect the energy transduction, reactive oxygen species (ROS) generation nervous system and heart are vulnerable – aging, degenerative disorders, and cancer.
Three mitochondrial cell death pathways Apoptosis inducing factor (AIF) Apoptotic protease activating factor-1(Apaf-1) Programmed cell death Cysteine protease family mtPTP (mitochondrial permeability transition pore) PS
Proton gradient– a central interconvertible currency of free energy (Mito. inner membrane)
P:O ratio: the number of molecules of inorganic phosphate incorporated into organic form per atom of oxygen consumed. the number of molecules of ATP synthesized per pair of electrons carried through electron transport. ATP synthesis: is quantitative as phosphate uptake, conversion of orthophosphate to organic phosphates. Electron pairs: are quantitative as oxygen uptake. matrix NADH: 2.5 matrix FADH2: 1.5 + 2,4-dinitrophenol: P/O ratio from 2.5 0 Ex. 6
96C 96T 192
97T 97C
98T 98C