Download

1 / 1
10 likes | 130 Views
V3 Loop Binding to CCR5 and CXCR4 of Rapid and Non Progressor HIV Patients in Baltimore MD. Background. V3 loop of gp120. CCR5 and CXCR4 Receptor.

E N D
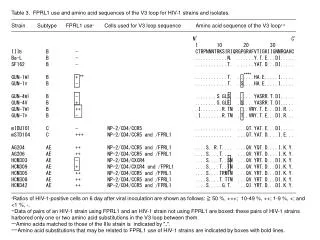
V3 Loop Binding to CCR5 and CXCR4 of Rapid and Non Progressor HIV Patients in Baltimore MD Background V3 loop of gp120 CCR5 and CXCR4 Receptor The HIV virus is a growing epidemic in the world that is responsible for the cause of death of millions of individuals. Modern medicine has not has found a cure and the high mutations that occur make the virus even harder to suppress once inside the body. The HIV virus is made of to key domains: the inner core and viral membrane. The purpose of this poster was to follow point mutations that occur in the V3 loop on the gp120 and CD4 counts of 2 needle using patients in Baltimore MD who have contracted the HIV virus. A non-progressor, the CD4 count did not drop throughout the study, and a rapid-progressor, the CD4 count dropped drastically through the study, patients V3 loop of the gp120 on the viral membrane were studied to find a reason as to why one patient died from HIV and the other did not die. The V3 loop binds to CCR5 and CXCR4 receptors found on the T-cell in the body. Research has shown that HIV that can utilize the V3 loop to bind CCR5 and CXCR4 is the most lethal combination for the destruction of the host T-cells. The V3 loop binds to the receptors through specific and nonspecific binding techniques and allows for the virus to enter into the T-cell leading to death of that T-cell. In this study it was possible to see as to why the rapid-progressor did indeed die from the actions of the HIV virus where as the non-progressor lived with a high CD4 count. These reasons all relate back to point mutations along the V3 loop and the structural alignment between the V3 loop when interacting with the CCR5 and CXCR4 receptors. Figure 1: Ribbon Diagram of V3 Loop; On the left is the conformation of the V3 loop in water. The Right picture show the conformation of the V3 loop in a polar solvent mimicking the actual binding conformation. The V3 loop is found on the gp120 protein and has a specific 3 dimensional structure that facilitates the HIV virus to binding to CCR5 and CXCR4 receptors on T-cells. There are two different sequences of the V3 loop: R5 and X4. For the purposes of this poster the R5 will only be considered. The V3 loop consists of 36 amino acids and has 2 stem and 1 crown domain as seen in the sequences bellow. The V3 loop begins with a disulfide bond between C1 and C36. The N-terminal stem consist of a conserved N-terminal loop and a small section of the β strand. The C-terminal stem is a conserved loose α helix. The crown region consists of a variable β strand, a conserved β hairpin turn, and a variable ridged loop. The β hairpin turn contains a highly conserved GPG sequence that is seen in all HIV isomers. The stem regions are believed to bind to the N-terminal domains of the CCR5 and CXCR4 receptors through salt bridges and nonspecific binding. The crown region of the V3 loop binds more specifically to the ECL2 of CCR5 and CXCR4 receptors utilizing the variable regions surrounding the β hairpin turn motif. Research has shown that the variable regions can undergo rapid point mutations that favor the binding of CCR5 or CXCR4. The ribbon diagram above show the conformation change that the V3 loop undergoes when it is in a water solution (Left) to a more polar solution (Right) that is typically seen upon binding to the different receptors. The correct 3 dimensional structure is necessary for aligning the crown region to the ECL2 allowing for viral binding. Figure 2: Sequence and location of the amino acids for CCR5 The CCR5 and CXCR4 7 transmembrane receptors that are utilized by the V3 loop of the gp120 to bind the HIV virus to the T-cell. The extra cellular N-terminal domain is interacts with the stem regions of the gp120 V3 loop and nonspecific binding from the β19 strand of the gp120. The binding of the β19 strand is specific for CCR5 recognition. Research has shown that S11 is specific for CCR5 biding where as R11 is specific for CXCR4 binding. There are many additional intermolecular interactions with the V3 loop (R9, R18, F20, GPG motif, and Y21) that cause recognition and binding to the receptors from the gp120 V3 motif. The CCR5 has an overall hydrophilic N-terminal and ECL2. Although within the ECL2 there is an area of 5 amino acids, KEGLH, which are hydrophobic and polar located towards the membrane where the β hairpin turn may interact. There are different interactions employed by the CXCR4 receptor. The CXCR4 receptor has 3 aspartic acids that interact with the β hairpin loop creating a hydrophobic pocket for the F20 to interact. The two receptors are very similar in function and structure although there are subtle differences that allow the V3 loop to be specific to either the CCR5 or CXCR4 receptor. Rapid-progressor Patient 10 Patient 10 was considered to be a rapid-progressor in the study done in Baltimore MD. This patient appears to have survived to the end of the study although with an extremely low CD4 count of 15 on visit 6. Looking at the CD4 count of this patient between visit 2 and visit 3 there was a significant drop in the patients CD4 count. The overall sequences of visits 1 and 2 were combined as well as the sequences from visits 3 and 4 which all showed similar V3 sequences. Comparing the sequences from visit 1&2 to the sequence from 3&4 there are point mutations that occur that shift the binding affinity from the CCR5 receptor the favor binding to the CXCR4 receptor. Using the point mutation study, substation of amino acids R11, R18, F20, and R30 showed that these amino acids affected the binding to CXCR4 but not the binding to CCR5. The amino acids that are conserved between visits 1-4 were R30 and R18. Therefore these amino acids could not have been the cause for such a great drop in the CD4 count between visits 2 and 3. Although the sequence did have a point mutation from F20 to L20. Both amino acids are hydrophobic and would fit into the hydrophobic pocket on the ELC2 loop. Since the L20 is smaller than the F20 it is possible to postulate that the binding affinity could be greater due to less steric hindrance. The second and most profound mutation that occurred was the change from S11 to R11. Research has shown that the serine is preferred in the CCR5 binding where as the arginine is preferred do to polar binding with other negatively charged amino acids in the ECL2 of the CXCR4 receptor. The change in affinity from the CCR5 to the CXCR4 has shown to be more lethal to HIV patients as apposed to binding affinity to CCR5 solely. This is a possible explanation as to why patient 10 was a rapid-progressor. Non-progressor Patient 12 Patient 12 was considered to be a non-progressor in the study done in Baltimore MD. This patient was considered a non-progressor by not observing a CD4 count drop throughout the duration of the study. After transcribing all of the mutant gp120 V3 loops that were obtained from the study, there were no point mutations that occurred within the overall sequences. For the purposes of this poster the V3 loop was examined. The amino acid sequence closely resembled that of the R5 sequence. Looking closely at the sequence, the selected amino acids appear to be specific for CCR5 binding and not CXCR4 binding. Looking at the point mutation study done in figure 4 the amino acids that appear to significantly affect the binding of V3 loop to CCR5 are still conserved (R11 and I26). Although a point mutation that is present throughout the entire study for this patient is a change from H13 to a P13. This amino acid is located in the middle of a β strand. The phi and psi angles needed to form a correct β strand are not able to be obtained with a P13 mutation. Therefore the 3 dimensional structure of the V3 loop is not oriented correctly with the ECL2 and crown region of the V3 loop, thus not allowing the binding to both the CCR5 and CXCR4. The CD4 count would not be affected due to the lack of the V3 loop to bind to the 2 receptors. Figure 3: The amino acid sequence of the overall mutational changes that occurred in patient 12 and patient 10. The 3 domains and secondary structures are coordinated on the figure with the colored bars. Figure 4: A point mutation study performed on the V3 loop measuring the binding affinities to the CCR5 and CXCR4 receptors. References Markham, Richard (1998) Proc. Natl. Acad. Sci.95, 12568-12573. Catasti, Paolo (1995) J. Biol. Chem.270, 2224-2232. Cormier, Emmanuel (2002) J. Virol.76, 8953-8957. Siciliano, Salvatore (1999) J. Biol. Chem.274, 1905-1913. Basmaciogullari , Stephane (2002) J. Viol.76, 10791-10800. Suphaphiphat, Pirada (2003) J. Virol.77, 3832-3837. Cilliers, Tonie (2003) J. Virol.77, 4449-4456. Tom Conrad Maugans Seegar Ursinus College Collegeville PA 19426