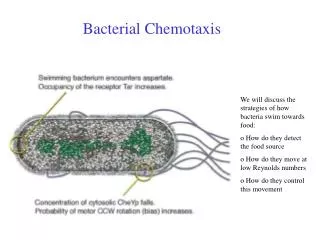
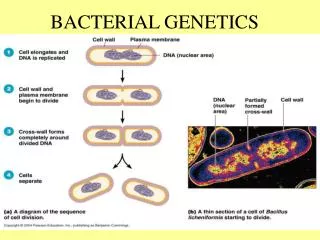
Bacterial pili
Bacterial pili. Pili in Gram-Negative Bacteria. Assembly of P-pilus in Gram negative bacteria.
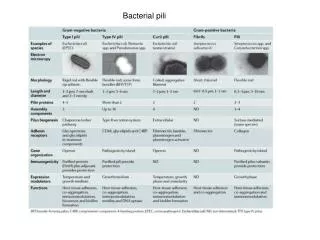

Bacterial pili
E N D
Presentation Transcript
Pili in Gram-Negative Bacteria
Assembly of P-pilus in Gram negative bacteria Fiber assembly by the chaperone–usher pathway. A schematic of P pilus assembly is shown. Single letters indicate the corresponding Pap protein. Fiber subunits enter the bacterial periplasm via the Sec system (YEG). In absence of the chaperone, subunits misfold, aggregate, and are proteolytically degraded. The chaperone (PapD) forms a soluble complex with each subunit (including the PapG adhesin, PapF, PapE, PapK, and PapA). The chaperone facilitates the folding of the subunit, stabilizes it, and caps its interactive surfaces while priming it for polymerization. Chaperone–subunit complexes are targeted to the pore-forming outer-membrane usher (PapC), where chaperone dissociation permits subunit incorporation into the growing fiber. The pilus adopts its quaternary structure outside the ce
Mechanism for Type IV pilus assembly recalls Type II secretion systems
Pili in Gram positive Bacteria • a | In the first step, proteins that contain the amino-acid motif LPXTG (where X denotes any amino acid) are targeted to the cell membrane by Sec-dependent secretion (not shown). This is followed by a sortase-mediated reaction (indicated by the arrows) in which the LPXTG motif is cleaved between the threonine (T) and glycine (G) residues. b | The reaction leads to the formation of an acyl-enzyme intermediate in which a covalent thioester bond is formed between the thiol group of a cysteine residue in the sortase and the carboxyl group of the pilin threonine residue. c | Oligomerization occurs after the nucleophilic attack provided by the -amino group of the lysine residue in the pilin motif on the cysteine residue of the sortase. d | The thioester bond between the pilin subunit and the sortase is targeted by the amino group of the pentapeptide of lipid II, the precursor of peptidoglycan. e | This leads to the formation of a membrane-associated, covalently linked, elongated pilus. NAG, N-acetyl glucosamine; NAM, N-acetyl muramic acid.
Gram negative Gram positive
Advances in understanding bacterial outer-membrane biogenesis: After synthesis at the inner leaflet of the inner membrane (IM), both lipopolysaccharide (LPS) and phospholipids (PL) are flipped to the outer leaflet of the IM. MsbA is required for the translocation of LPS and possibly phospholipids across the IM, but helical transmembrane domains have also been shown to translocate phospholipids. How both LPS and phospholipids travel from the IM to the outer membrane (OM), and how LPS is flipped to its outer leaflet, remains unknown, although the OM protein (OMP) Imp is required for the assembly of LPS. Both OM lipoproteins and OMPs are synthesized in the cytoplasm and are translocated across the IM by the Sec translocon. After undergoing lipid modification and signal-sequence processing (not shown), OM lipoproteins interact with the ATP-binding cassette (ABC) transporter LolCDE, which hands them over to the periplasmic chaperone LolA. LolA escorts OM lipoproteins across the periplasm and delivers them to the OM-assembly site, the OM lipoprotein LolB. After translocation through the Sec machinery, OMPs are transported to the OM by an unknown mechanism, although the periplasmic chaperones Skp, DegP and SurA have been implicated. At the OM, the YaeT/YfgL/YfiO/NlpB complex assembles OMPs by an unknown mechanism.
Sorting and outer membrane localization of lipoproteins by the Lol system. “In” and “Out” represent inner membrane-specific and outer membrane-specific lipoproteins, respectively. The ABC transporter LolCDE recognizes outer membrane-directed lipoproteins and releases them from the inner membrane, causing the formation of a complex between one molecule each of lipoprotein and LolA, a periplasmic carrier protein. The LolA-lipoprotein complex traverses the periplasmic space and lipoproteins are transferred to the outer membrane receptor protein LolB, followed by incorporation into the outer membrane. LolB is itself an outer membrane lipoprotein. Inner membrane-specific lipoproteins with Asp at position 2 avoid recognition by LolCDE and thus remain anchored to the inner membrane.
The lipoprotein-releasing apparatus. (A) Reconstitution of lipoprotein-releasing activity into proteoliposomes. Inner membrane proteins are solubilized with a detergent and then reconstituted into proteoliposomes together with purified lipoproteins and E. coli phospholipids. Proteoliposomes are frozen, thawed and sonicated to load ATP, followed by incubation with LolA. Lipoprotein-releasing activity is measured as the amount of lipoproteins recovered in the supernatant after ultracentrifugation. (B) The LolCDE complex conferring the lipoprotein-releasing activity. The membrane topology of LolC and LolE is based on topology prediction and biochemical analyses. (C) The LolCDE avoidance function of Asp depends on phospholipids. Proteoliposomes are reconstituted with purified LolCDE, E. coli phospholipids (PL) and Pal or Pal(S2D). Inner membrane-specific Pal(S2D) possessing Asp in place of Ser at position 2 is not released from proteoliposomes reconstituted with PL. P and S indicate the pellet and supernatant fractions, respectively.
MICRB311 Problem 18 Several lipoproteins have been identified by biochemical methods in both, the inner and the outer membrane of Gretzkyrinia oileriensis. Genes homologous to LolABCDE were identified in this organism. The genes are called GolABCDE in G oileriensis. To understand how the lipoproteins are sorted to the inner membrane and the outer membrane in this special bacterium, Dr. Clonit purified GolCBE proteins and reconstituted them in vesicles containing phospholipids and the lipoprotein LipZ. LipZ is a lipoprotein normally found in the outer membrane of G. oileriensis. Dr Right incubated the vesicles with and without ATP and GolA, then centrifuged the samples and analyzed the levels of LipZ in the pellet and the supernatant by Western blot, using antibodies directed against LipZ. These are the results: + GolA GolA - A ATP - ATP - ATP + ATP + Dr Clonit was very surprised when discovered that the amino acid in position 2 of the sequence of mature LipZ was an aspartic acid (D, negatively charged amino acid). He mutagenized this D residue to three different amino acids: Serine (hydrophilic, S); L (leucine, hydrophobic) and R (arginine, positively charged). Then he repeated the experiment using these mutants. These are the results obtained. B D2S D2L D2R • Explain the results shown in the figure A. What would be the role for the different Gol proteins included in this experiment. Is ATP needed for GolA activity? If you answer is yes, explain why. • What is the role for the amino acid in position 2? According to the experiment presented in figure 2, in which compartment would you expect to find lipoproteins containing a D residue at position 2 in G. olieriensis? What about lipoproteins containing an S in that position? And if the amino acid is an R?