The number of B ’s born ~ the number of A-B pairs: ~ a ( x,t ) b ( x,t ) l d t
440 likes | 815 Views
a ( x,t ) = the number of A’ s ; b ( x,t )= the number of B ‘s The number of B ’s dying ~ proportional to their number ~ - m b ( x,t ) d t The number of B ’s born ~ the number of A-B pairs: ~ a ( x,t ) b ( x,t ) l d t In total:

The number of B ’s born ~ the number of A-B pairs: ~ a ( x,t ) b ( x,t ) l d t
E N D
Presentation Transcript
a ( x,t ) = the number of A’s ; b ( x,t )= the number of B ‘s The number of B’s dying~ proportional to their number ~ - m b ( x,t )d t The number of B’s born~the number of A-B pairs: ~ a ( x,t ) b ( x,t ) l d t In total: d b ( x,t ) ~ ( a ( x,t ) l - m )b ( x,t ) d t+diffusion Continuum d t 0: b. = ( a l- m)b + Db D b a. = Da D a
Since the classical works of Malthus and Verhulst it has been recognized that most of the processes in living systems are autocatalytic and thus are characterized by exponential growth. In fact, the appearance of an autocatalytic molecule may be considered as the origin of life. Autocatalytic system are shown to admit self organization in the presence of fluctuating environment. The exponential amplification of ``good'' fluctuations in the catalysis parameters prevails, in the situations discussed above, the globally hostile environment and is robust against the random motion of both the reactants and the catalysts. Our result may be interpreted as an indication that ``life'' (in the above sense) is resilient and is able to adapt itself to the changing environment. The applicability of this model ranges from biological evolution to the role of enzymes in chemical reactions and even in social or financial settings.
Since Malthus and Verhulst: • autocatalytic and thus • exponential growth. • Autocatalytic system admit self organization in the presence of fluctuating environment. Exponential amplification of ``good'' fluctuations in the catalysis parameters • prevails, the globally hostile environment and • is robust against the random motion of both the reactants and the catalysts. • The applicability of this model ranges from biological evolution to the role of enzymes in chemical reactions and even in social or financial settings.
Since Malthus and Verhulst: • autocatalytic and thus • exponential growth. • Autocatalytic system admit self organization in the presence of fluctuating environment. Exponential amplification of ``good'' fluctuations in the catalysis parameters • prevails, the globally hostile environment and • is robust against the random motion of both the reactants and the catalysts. • The applicability of this model ranges from biological evolution to the role of enzymes in chemical reactions and even in social or financial settings.
Malthus exponential growth when <birth> > <death > b.(t)= a b(t) => exponential increase for a > 0 Here Super-Malthus: always exponential growth:Natural death mb(x,t)does not lead to saturation. For very largeb(x,t)’s competition necessary (Verhuulst 1838): Logistic equationb.(t)= a b(t)– g b(t)2 Montroll: “almost all the social phenomena, except in their relatively brief abnormal times obey the logistic growth''. Lotka and Volterra: b(t)= the size of an animal/plant population, a = the aggregated effects of birth and natural death, -g b(t)2 = competition for limited resources.
Malthus exponential growth when <birth> > <death > b.(t)= a b(t) => exponential increase for a > 0 Here Super-Malthus: always exponential growth:Natural death mb(x,t)does not lead to saturation. For very largeb(x,t)’s competition necessary (Verhuulst 1838): Logistic equationb.(t)= a b(t)– g b(t)2 Montroll: “almost all the social phenomena, except in their relatively brief abnormal times obey the logistic growth''. Lotka and Volterra: b(t)= the size of an animal/plant population, a = the aggregated effects of birth and natural death, -g b(t)2 = competition for limited resources.
Economics, Aoki - b(t) = the total product demand in a market. -a b(t) = emergence of new products ~ size of the market. -g b(t)2 the products have to compete with one another within a finite total potential market. Finance, Solomon and Levy have suggested that - b(t) = the total capital within a financial system. -a b(t) = the average returns that the system offers, -g b(t)2 the effects of competition and
Economics, Aoki - b(t) = the total product demand in a market. -a b(t) = emergence of new products ~ size of the market. -g b(t)2 the products have to compete with one another within a finite total potential market. Finance, Solomon and Levy have suggested that - b(t) = the total capital within a financial system. -a b(t) = the average returns that the system offers, -g b(t)2 the effects of competition and
Local Competition Many of the mechanisms limiting the B proliferation can be described as competition. Competition for a resource means that whenever 2 B agents need the resource either to survive or to proliferate only one of them will get it and the other will die (or fail to proliferate). In ecology competition can be over food or water, while in the immune system competition can be over access to an antigen. Such a system is sometimes called a Lotka Voltera system [18, 19] and can be classically described using a local differential equation: b.(x,t)= (la(x,t)- m)b(x,t)– g b(x,t)2 + Db D b(x,t) If (la(x,t)– m) < 0 The adaptive islands appear here too and increase greatly the sustainability of the B population. However, the height of the B islands is finite and for discrete B’s and finite volume the population will eventually die by some cataclysm which brings it to the b=0 fixed point.
Local Competition Lotka Voltera system (use PDE notation to symbolize particles interactions in spite of its inadequacy as dynamical predictor): b.(x,t)= (la(x,t)- m)b(x,t)– g b(x,t)2 + Db D b(x,t)If (la(x,t)– m) < 0 Adaptive islands appear here too and increase greatly the sustainability of the B population. However, the height of the B islands is finite and for discrete B’s and finite volume the population will eventually die by some cataclysm which brings it to the b=0 fixed point.
Global (long range/“mean field !?”) Competition The globally averaged version of AB system with B competition (logistic, Lotka-Volterra system):b.(t)= a b(t)– g b(t)2where b(t) = < b(x,t) >x The linear term a b(t)can be obtained by averaging over the spatial locations x in: b.(x,t)= (l a(x,t)- m )b(x,t)– g b(x,t)2 + Db D b(x,t)and denotinga= (l a(t)- m ) the termb(t)2 = < b(x,t) >x2cannot be the result of averaging over local (nonlinear) terms:b2 (t) = < b(x,t) >x2 < b(x,t)2 >x
Nonlinear global terms =averages over infinite range interactions:b(t)2 = < b(x,t) >x2 = < b(x,t) <b(x,t)>x >x , mean field! b.(x,t)=(la(x,t)-m )b(x,t)–g b(x,t) <b(x,t)>x+ Db D b(x,t)Stable state (neglect diffusion): b.(x,t)=0=[(la(x,t)-m )– g<b(x,t)>x] b(x,t) Except (measure 0 set of x’s) where(l a(x,t) - m ) - g<b(x,t)> x =0 => localization:for almost all xb(x,t) = 0 In fact for (l a(x,t) - m ) - g<b(x,t)>x> 0, b(x,t) = 0 is unstable (b.(x,t) >0 if b(x,0) =e >0) => b(x,t) evolves until: <b(x,t)> x = (l a(xmax,t) - m )The entire B population is an island aroundxmaxwith populationb(xmax,t) = (l a(xmax,t) - m ) x Volume
Rather they can appear as averages over infinite range interactions:b(t)2 = < b(x,t) >x2 = < b(x,t)b(t) >x , mean field => greatly enhancing the localization effects b.(x,t)= (la(x,t)- m )b(x,t)– g b(x,t) <b(t)>+ Db D b(x,t)Fixed point: b.(x,t) =0 => (neglect diffusion) (l a(x,t)- m ) = g<b(t)> or b(x,t) = 0 I.e b vanishes in the entire space except the singular locations where a(x,t) takes a specific value (which turns out to be its maximum). The entire B population is concentrated in one island.The continuum limit is NEVER correct.(even for (l a0- m > 0)
Globalization of Competition => Localization of Growth The linear term a b(t)can be obtained by averaging over the spatial locations x in: b.(x,t)= (l a(x,t)- m )b(x,t)– g b(x,t)2 + Db D b(x,t) the nonlinear termg b(t)2cannot bethe result of averaging overlocal (nonlinear) terms:b2 (t) = < b(x,t)>x2 < b(x,t)2 >x Global Relevant: O2/O3, greenhouse, polution, inflation,capital The globally averaged version of AB system with B competition (logistic, Lotka-Volterra, Verhuulst 1838/Malthus):b.(t)= a b(t)– g b(t)2where b(t) = < b(x,t) >x
Finite Competition Range R b.(x,t)=(la(x,t)- m )b(x,t)– gb(x,t)<b(x,t)>R+ Db D b(x,t) <b(x,t)>R= S|y-x-1|<Rb(y,t) / S|y-x-1|<R1
The necessity for B to find a way to the new A pile emphasizes the difference between - random (space time-uncorrelated) noise – where A’s would disappear from one site and would reappear in another randomly uncorrelated site. • and • diffusive (space-time correlated) noise – where A just jumps between neighboring sites and allows (descendents of) the B’s to follow them. Continuityof the random hostile environment in time is a crucial feature for adaptability and life emergence.
Saturation Effects; Globalization of Competition leads to Localization of Growth
Local Competition One spatially extended generalization of the Lotka Voltera system: b.(x,t)= (la(x,t)- m)b(x,t)– g b(x,t)2 + Db D b(x,t) a(x,t) is not a continuum function: it is a stochastic integer at each site x and it changes in time according to the jumping diffusion rules discussed before. If (la(x,t)– m) < 0 => localized adaptive islands of finite hight. - Improved life expectation but b=0 fixed point for discrete B.
The Local Competition equation b.(x,t)= (l a(x,t)- m )b(x,t)– g b(x,t)2 + Db D b(x,t) is not the natural space extended version ofb.(t)= a b(t)– g b(t)2 . Indeed averaging <g b(x,t)2 > does notgive g b(t)2g< b(x,t) > 2 average overthe nonlocal (infinite range) term < g b(x,t) <b(x,t)>x >x = g< b(x,t) >x2does. One is lead to the global competition equation:b.(x,t)= (l a(x,t)- m )b(x,t)-gb(x,t)<b(x,t)>x+DbDb(x,t)
b.(x,t)=(la(x,t)-m )b(x,t)–g b(x,t) <b(x,t)>x+ Db D b(x,t)Stable state (neglecting diffusion) is given by : b.(x,t)=0=[(l a(x,t)-m )– g<b(x,t)>x]b(x,t) (1) Concentrate on the site xmax with the property:a(xmax,t) > a(x,t): x xmaxaccording (1) its population keeps increasing until[(l a(xmax,t) m )– g<b(x,t)>x] = 0 (2) This fixes <b(x,t)>x . But this means that for x xmax[(l a(x,t) m )– g<b(x,t)>x]< 0 And according (1)b(x,t) will decrease until b(x,t) =0 x xmax The entireBpopulation is in an island aroundxmaxwith populationb(xmax,t) ~ (l a(xmax,t) - m ) x Volume
Behavioral Finance B= financial traders, Patches= "herding" behavior (despite the fact that we do not introduce communicationbetween B’s).. • May explain some paradoxes in finance E.g • between theefficient market hypothesis (absence of systematic profit opportunitiesin equilibrium markets) and • the actual profits that investorsextract daily from themarket.
Saturation Effects; Globalization of Competition leads to Localization of Growth
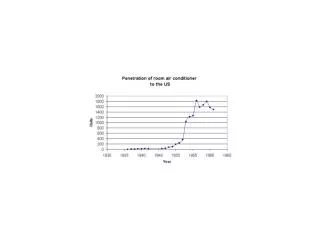
Malthus exponential growth when <birth> > <death > b.(t)= a b(t) => exponential increase for a > 0 Here Super-Malthus: always exponential growth:Natural death mb(x,t)does not lead to saturation. For very largeb(x,t)’s competition necessary (Verhuulst 1838): Logistic equationb.(t)= a b(t)– g b(t)2 Montroll: “almost all the social phenomena, except in their relatively brief abnormal times obey the logistic growth''. Tasmania Sheep Lotka and Volterra: b(t)= the size of an animal/plant population, a = the aggregated effects of birth and natural death, -g b(t)2 = competition for limited resources.
Economics, Aoki - b(t) = the total product demand in a market. -a b(t) = emergence of new products ~ size of the market. -g b(t)2 the products have to compete with one another within a finite total potential market. Finance, Solomon and Levy have suggested that - b(t) = the total capital within a financial system. -a b(t) = the average returns that the system offers, -g b(t)2 the effects of competition and
Local Competition One spatially extended generalization of the Lotka Voltera system: b.(x,t)= (la(x,t)- m)b(x,t)– g b(x,t)2 + Db D b(x,t) a(x,t) is not a continuum function: it is a stochastic integer at each site x and it changes in time according to the jumping diffusion rules discussed before. If (la(x,t)– m) < 0 => localized adaptive islands of finite hight. - Improved life expectation but b=0 fixed point for discrete B.
The Local Competition equation b.(x,t)= (l a(x,t)- m )b(x,t)– g b(x,t)2 + Db D b(x,t) is not the natural space extended version ofb.(t)= a b(t)– g b(t)2 . Indeed averaging <g b(x,t)2 > does notgive g b(t)2g< b(x,t) > 2 average overthe nonlocal (infinite range) term < g b(x,t) <b(x,t)>x >x = g< b(x,t) >x2does. One is lead to the global competition equation:b.(x,t)= (l a(x,t)- m )b(x,t)-gb(x,t)<b(x,t)>x+DbDb(x,t)
b.(x,t)=(la(x,t)-m )b(x,t)–g b(x,t) <b(x,t)>x+ Db D b(x,t)Stable state (neglecting diffusion) is given by : b.(x,t)=0=[(l a(x,t)-m )– g<b(x,t)>x]b(x,t) (1) Concentrate on the site xmax with the property:a(xmax,t) > a(x,t): x xmaxaccording (1) its population keeps increasing until[(l a(xmax,t) -m )– g<b(x,t)>x] = 0 (2) This fixes <b(x,t)>x . But this means that for x xmax[(l a(x,t) -m )– g<b(x,t)>x]< 0 And according (1)b(x,t) will decrease until b(x,t) =0 x xmax The entireBpopulation is in an island aroundxmaxwith populationb(xmax,t) ~ (l a(xmax,t) - m ) x Volume
(m-l a0) l