Download
1 / 38
390 likes | 1.01k Views
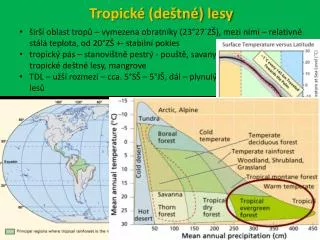
Tropické (deštné) lesy. š irší oblast tropů – vymezena obratníky (23°27´ZŠ), mezi nimi – relativně stálá teplota, od 20°ZŠ +- stabilní pokles t ropický pás – stanovištně pestrý - pouště, savany, opadavé tropické lesy, tropické deštné lesy, mangrove

E N D
Tropické (deštné) lesy • širší oblast tropů – vymezena obratníky (23°27´ZŠ), mezi nimi – relativně stálá teplota, od 20°ZŠ +- stabilní pokles • tropický pás – stanovištně pestrý - pouště, savany, opadavé tropické lesy, tropické deštné lesy, mangrove • TDL – užší rozmezí – cca. 5°SŠ – 5°JŠ, dál – plynulý přechod do sezónních lesů
Tropické klima • klima dáno zejména ročním a denním chodem teploty a množstvím a rozložením srážek (dáno ZŠ a nm. výškou) – vše determinuje množství slunečního záření • Poblíž rovníku - průměrná měsíční teplota neklesá pod 25°C + stálá délka dne (12 h) během roku • rozdíly v teplotě dne a noci – větší než sezónní • vliv nehomogenního přísunu sluneční energie na Zemi (ZŠ a náklon zemské osy+oběh Z kolem S) • rovník – sluneční paprsky – kolmo – největší příkon, neměnný; dál k obratníkům – pořád dost „kolmo“ (max. odklon od kolmého směru 23°27´) • k pólům – čím dál šikměji – proporčně míň energie na stejnou plochu, navíc – ztráty v atmosféře (vliv reliéfu – J svah 10° - ozářenost stejná jako na rovině o 10° blíž k rovníku) • od rovníku na S a J – sezónní změna délky fotoperiody
Tropické klima • nestejné prohřívání různých ZŠ + odlišná tepelná kapacita moře a pevniny – nehomogenity – zásadní pro vznik a průběh tlakových útvarů – determinují klima konkrétní oblasti • tlakové níže (ohřátý vzduch stoupá) x tlakové výše – místa poklesu studených vzdušných mas – horizontální proudění po gradientu tlaku – vítr • pravidelnost v teplotním režimu – pravidelné větrné systémy
Tropické klima • kolem rovníku zóna trvalých tlakových níží – ITCZ (Intertropicalconvergencezone)..PROČ? Trvalé stoupání ohřívajícího se vzduchu – pokles tlaku • největší ohřev – přes den stoupá ohřátý vlhký vzduch – nahoře ochlazení – kondenzace – lokální odpolední srážky • už vypršené vzdušné masy – ve výšce stáčení na S a J (antipasáty), klesají k povrchu kolem obratníků (tl. výše-koňské šířky)– „nasávají vlhkost“ – proto zóna pouští • návrat k rovníku (do oblastí nízkého tl.), stáčení vlivem Coriolisovy síly = suché a teplé pasáty (tradewinds)
Tropické klima • tato cirkulace – potlačení/zesílení vlivem odlišného rozložení moří a pevnin – sezónní tlakové útvary + navazující proudění – nejznámější – monzuny (ale taky mistrál, blizard, fén…) • atmosférická cirkulace – spojené nádoby s cirkulací mořskou (vliv odlišné teploty a salinity vody)
Tropické klima • sezonalita tropů tedy dána množstvím a rozložením srážek - na rovníku – perhumidní oblast, na S a J – zvýrazňují se období sucha (i poblíž rovníku – nepatrná maxima srážek v období rovnodenností – kulminace slunce nad rovníkem) • dál – 2/1 období dešťů – podle ročních posunů ITCZ, zhruba od 5°ZŠ – vliv suchých pasátů – sezónní opadavé lesy x JV Asie – větší rozsah tropů dán monzuny • roční úhrn srážek na rovníku >3000 mm, ale i větší výpar (> 1850 mm) Max. srážek/rok – 26 461 mm – Cherrapunji, Indie Max./den – 1870 mm ostrov Reunion
Mikroklima TDL • makroklima – stabilní během roku x mikroklima – fluktuace během dne • přehřívání povrchu vegetace přes den (=horní část canopy) a ochlazování v noci x interiér lesa (stabilní teplota) • obdobně – chod vlhkosti => odlišné adaptace druhů obou mikrostanovišť • tuhé, lesklé, xeromorfní listy • silná kutikula, i zanořené průduchy • často druhy pokojových rostlin, snášející nízkou vzdušnou vlhkost (Ficus, Anthurium…) • kolísání teploty až o 15°C, vlhkosti 100-60% • sciofilní druhy – tenké, široké, mnohdy členité listy – kapradiny, vranečky • kapací špičky (drip-tips) • pod 1% slunečního záření – absence zelených rostlin (saprotrofní či parazitické druhy –Thismia, Raflesia..) • hojné saprotrofní houby
Půdy TDL • Obecné paradigma: • tropické půdy – staré (až třetihory) – současně perhumidní oblast - > silně zvětralé a ochuzené o živiny, často kyselé (lateritické zvětrávání – vymývání silikátů, ale i bází – zůstávají oxidy železa a hliníku, podzolizace – vymývání jílových částic, huminovýchkys, bází…nízké pH) • většina živin – vázána v biomase • vlivem klimatu – rychlá dekompozice a krátké cykly prvků • minimum organických látek v půdě
Půdy TDL • platí často, ale NE VŽDY (viz várzea – Amazonie – inundační zóna „whitewaters“, mladší vulkanické substráty – Indonésie, PNG – bohaté na živiny) …vers. igapó („blackwaters“) • viz rozložení živin mezi nadzemní biomasou a půdou – specifické pro konkrétní živiny a pro různé části biomu várzeaforest igapoforest
Půdy TDL – role mykorhizy • oproti původním předpokladům (viz nejzásadnější role mykorhiz v boreálních eks.) – velmi hojné mykorhizy i v TDL (odhad 70-90% všech stromů) • Ektomykorhiza (paleotropy – Dipterocarpaceae, Fagacea, častá v půdách s pomalejším koloběhem C a malým obsahem N – horské lesy, druhově specifičtější interakce, častá u dominant) • Arbuskulární m. (=endomykorhiza; v neotropech; arbuskuly = stromečkovité útvary hyf větvících se uvnitř buněk kořene rostliny, půdy s rychlým turn-over, nedostatkem P) • mnoho výhod M – snazší „vytahování“ P a N z půd, zejména z biomasy – možná i rychlejší uvolňování org. P • pionýrské stromy – lépe klíčí při infekci AM (vliv dodatku P) • vyšší rezistence x toxickým kovům (Al, Fe), ale i k napadení fytopatogeny a k suchu – hlavně ektomykorhiza
Vegetační struktura TDL • V jednotlivých oblastech – podobné zastoupení životních forem, ty ale reprezentovány často odlišnými taxony
Vegetační struktura TDL • Stromy • vertikální stratifikace – 2-3 patra, částečně přecházejí jedno v druhé • nejvyšší – tzv. emergents (nesouvislý zápoj, až 60 m, široké koruny – nemají konkurenci o světlo) • střední canopylayer – 25-35 m, zapojená vrstva, koruny menší, kulatější • spodní korunové patro – 10-15 m, ne zcela souvislé, koruny spíš užší – vychytávají sporadicky prostupující světlo
Vegetační struktura TDL - stromy • Kmeny štíhlé, větví se vysoko, tenká kůra; kořeny mělké + buttressroots – opora (deskovité x chůdovité kořeny) • málo vyvinuté bylinné patro – minimum světla projde až „na dno“, kompetice se stromy o živiny – minimum trav • obrovská diverzita stromů – základ celkové biodiverzity TDL • více než 400 druhů stromů/ha (Ekvádor) x i druhově chudé TDL (dvoukřídláčové lesy v Malajsii, lesy karibských ostrovů s dominancí Mora excelsaaj. • gapová dynamika – udržuje diverzitu (trade-off mezi r- a K-stratégy)
Vegetační struktura TDL 2. Liány • 90 % všech lián – tropy - dáno silnou kompeticí o světlo – epifyty a liány vyřešily problém výstavby kmene • Jak vyšplhat na slunce? Ovíjivé stonky, úponky (mučenky, Cissus, révovité), příčepivé kořeny (arónovité), opěrné trny – palmy (Arecaceae– např. ratan, r. Calamus) • více lián – neotropy x vyšší stromy – JV Asie – řada konsekvencí v adaptacích živočichů…
Vegetační struktura TDL – epifyty a hemiepifyty • Epifyty – nejčastěji bromélie (JA), orchideje (všude) a kapradiny (časté v JV Asii) • neškodí přímo (na rozdíl od parazitů a hemiparazitů), ale: nutrientpiracy, v případě velkého množství epifytů a lián – zatěžují nosný strom, náchylnější k polámání (mlžné horské lesy) • hl. adaptace epifytů – na nedostatek vody a živin (bromélie – fytotelmy, šupiny pro příjem vody, parožnatky speciální listy bez chlorofylu – k hromadění opadu, Asplenium – zachytává opad díky široce rostlým listům; adaptace na dočasné sucho – viz epifytické kaktusy (JA) • Hemiepifyty – známé škrtiče – hojně r. Ficus (klíčí jako epifyt, pak spouští kořeny na zem), některé palmy (Arecaceae)– klíčí na zemi, šplhají nahoru, pak spojení ztrácí
Fenologie a reprodukce rostlin TDL • chybí klimatická sezonalita – málo výrazná periodicita (občas autonomní periodicita daná populačními cykly druhů – viz Dipterocarpaceae– mast years– výhoda je přečíslení predátorů semen v čase, mezi druhy r. Shorea - nichepartitioning díky rozptýlení kvetení v čase - snížení kompetice o opylovače • výměna listů – n týdnů- n let, některé druhy shazují listí naráz, jiné vyměňují za nové – velká časoprostorová variabilita v rámci druhu i na 1 stromě, stejně tak i s kvetením a plozením!
Fenologie a reprodukce rostlin TDL • málo větrosprašných druhů (pionýři) x nejčastější opylovači – hmyz (častá specializace), nektarofágní ptáci a netopýři (kauliflorie, baziflorie) • koevoluce rostlin a opylovačů – hmyzosnubné rostliny – zářivé barvy, ne tak robustní květy x nektarofágní ptáci – často červené, robustní květy (Heliconia) x květy závislé na netopýrech – hustší nektar, otvírají se v noci, intenzívní pach, bílé… • i rozptyl semen – často živočichové (plodožraví kaloni a listonosové, aguti, ptáci, dokonce i ryby)
Koevoluce v TDL • extrémní příklady mutualistické koevoluce – listonos r. Glossophaga a liána r. Marcgravia, nebo fíky a „fíkové vosičky“, případně orchidej „hvězda Madagaskaru“ a lišaj Xanthopan morgani praedicta… • koevoluce i jako „armsrace“ – např. herbivořivers. rostliny (špatně stravitelná celulóza, chemická a mechanická obrana rostlin, mast-years…)
Konvergentní evoluce (v) TDL • vývoj podobných forem (vybavených podobnými adaptacemi) uvnitř navzájem nepříbuzných skupin - reflektuje podobné selekční vlivy prostředí • např. mnoho lián z nepříbuzných čeledí rostlin, • tukani (JA) x zoborožci (Afr, J-V Asie) – plodožraví obyvatelé dutin • rajky (PNG) x lemčíci (Austrálie) x kotingy a manakiny (JA) – behaviorální konvergence – extrémní namlouvací rituály – spojováno s neustálým dostatkem potravy, resp. nízkou intenzitou predace (proto chybí v JV Asii, Africe)
Fauna TDL • nejdiverzifikovanější ze všech biomů souše (druhy, rody, čeledi…) • ALE: biomasa velkých herbivorů je nižší než např. v savaně, stepi (nedostupnost potravy – většina v canopy, tam se ale zpravidla nedostanou (výjimka např. lenochodi, vřešťani…)
stratifikace (prostorové odlišení nik), potravní specializace (funkční odlišení) + časová segregace – dobrým příkladem všeho – ptáci (ráj v neotropech – zřejmě vliv stálého přísunu potravy (ovoce, semena, výhonky… x JV Asie – částečně cykly Dipterocarpacea)
SAVCI - druhově nejpočetnější primáti a letouni (až 50% druhů tropických savců!), ostatní – rel. málo, většina stromových • na zemi – nika sběračů plodů (kančilové x aguti x lesní antilopy+prasatovití), jen pár velkých predátorů (největší – kočkovité šelmy) x ALE!! - nejúspěšnější predátoři TDL – hadi!
odlišná struktura TDL (JA x JV Asie) – vedla k evoluci odlišných adaptací… …zpomalení pádu – klouzavý let vers. ovíjivé ocasy • maximum „plachtičů“ - JV Asie - 60 druhů plachtících obratlovců (žáby, gekoni, ještěři, poletuchy, letuchy, 1 druh hada) • únik predátorům, pohyb v korunách • Afrika 6 druhů, Amerika – jen pár druhů žab, ne let, spíš pád, jen málo zpomalený létavka černoblanná (Rhacophorusnigropalmatus) bojga Chrysopeleasp. poletucha velká (Petauristapetaurista) letucha malajská (Cynocephalusvariegatus) dráček Dracovolans
Vysvětlení odlišných adaptací stromových živočichů • 1. hypotéza vysokých stromů: pralesy JV Asie vyšší než kdekoliv jinde (Dipterocarpaceae) + průměrně nižší rychlost větru • 2. hypotéza přerušovaného lesa: korunové patro má málo lián → koruny nejsou propojené, jako v Africe a Americe (zde se vyvinul nezávisle na sobě u několika skupin chápavý ocas – stromoví mravenečníci, kynkažu, chápani…) • 3. teorie potravní pouště: v korunovém patře dominují Dipterocarpaceae,které jsou téměř nepoživatelné (toxiny) • → nutnost zvýšeného pohybu a vyhledávání zdrojů
Bezobratlí živočichové • řádově početnější než obratlovci (nejen v tropech) – cca. 1,8 mil. druhů, z toho cca. 1,3 mil. živočichů, z nich asi 900 000 – hmyz, z toho skoro 80% v tropech, velká část – herbivoři, saprofágové • herbivorie=„kořist“ sice neuteče, přesto dělá problémy: (ne)stravitelnost celulózy, poměr C:N, „obrana“ rostlin • významní herbivoři – termiti – symbióza • s brvitkami • značná část biomasy TDL – mravenci (významní herbivoři x predátoři) - koevoluce s houbami a rostlinami - zásadní predátoři, nekrofágové TDL - 12500 druhů, většina – v tropech
Opylovači • mnohé taxony, nejen hmyz (viz lokální dominance kaloňů) • větrosprašné rostliny – zřídka, dáno hl. velkými vzdálenostmi mezi stejnými stromy + mikroklimatem interiéru lesa) • generalisté i specialisté, motivace – nejen nektar, ale i pyl, hl. u hmyzu (mnoho bílkovin)
Konvergentní evoluce „obratlovčích“ opylovačů ve všech tropických oblastech – PROČ? • hl. výhoda – přenos velkého množství pylu na velkou vzdálenost (x hmyz) Nectariniidae (strdimilovití) Phyllostomidae (listonosovití) Pteropodidae (kaloňovití) Trochilidae (kolibříkovití) Psittacidae (papouškovití) Meliphagidae (kystráčkovití) Zosteropidae (kruhoočkovití)
Neotropy– úzká specializace obratlovčích opylovačů, více rodů opylováno obratlovci než v paleotropech • Paleotropy– nižší míra specializovanosti a frekvence výskytu, přitom Afrika je „na půl cesty“ mezi JA a JV Asií a Australasií PROČ? 1) Neotropy – rostliny plodí předvídatelněji, trvale dostupná potrava, nadto druhově pestrá – možnost užší specializace 2) evoluční omezení – např. některé skupiny – menší variabilita – nesnadná radiace 3) rostliny jednodušeji „přepnou“ z opylování hmyzem na opylování malými kolibříky, než např. těžkopádnými papoušky (viz nutná změna morfologie květu) x
Makroekologicképatternsbiodiverzity - společné trendy • Druhová rozmanitost se charakteristicky mění v latitudinálním a altitudinálním měřítku. • Druhová rozmanitost je kladně korelována se srážkami a průměrnou měsíční teplotou. • Druhová rozmanitost na regionální úrovni má vztah k rozmanitosti na lokální škále (a naopak).
Makroekologicképatternsbiodiverzity - společné trendy • Druhová rozmanitost se mění synergicky napříč taxonomicky různými skupinami organismů. • Velmi dobře druhovou rozmanitost predikuje produktivita a MDE. • Výše uvedenému odpovídají TDL jakožto místa s potenciálně nejvyšší druhovou rozmanitostí… • …a ono to tak opravdu je (což se snaží vysvětlit také celá řada dalších hypotéz…)
Ohrožení TDL • odhad původní rozlohy TDL asi 16 mil km2 • vr. 1982 pouze 9,5 mil. km2, během následující dekády zmizelo dalších 2,8 mil km2 • v současnosti ubývá kolem 140 tis. km2 ročně (rozloha bývalé ČSR) • zhruba polovina tohoto území je kompletně zničena, zbytek poškozen (např. výběrová těžba tropických dřev – mimo vytěžení části stromů – fragmentace) • velmi malá část územně chráněna • Příčiny ztrát rozlohy TDL • >50% - maloplošné „soukromé“ zemědělství – shiftingcultivation= slash and burn • v JA – po skončení pěstování plodin následuje převod na pastviny dobytka • těžba palivového dříví (zhruba 13% světové populace vaří na ohni) • komerční těžba – 21% ročních ztrát • cílené mýcení pro založení pastvin – cca. 11% • zhruba 10% - plantáže, infrastruktura
Primární les Tropický les převedený na plantáž palmy olejné Primární les po výběrové těžbě
Je třeba chránit nejintenzívněji právě TDL? • více přístupů k ochraně světové diverzity…viz biodiversity hot-spots – rozhodně většina ne leží právě v oblasti TDL!
Globální ztráty rozlohy biomů Mediterranean Forests Temperate Grasslands & Woodlands Temperate Broadleaf Forest Tropical Dry Forest Tropical Grasslands Tropical Coniferous Forest Tropical Moist Forest 0 50 100 Percent of habitat (biome) remaining Source: Millennium Ecosystem Assessment
Kontinentální step Mediteránní lesy a křoviny
Tropický deštný les a jeho ochranářská priorita? • Ochranářské priority je potřeba hledat bez primárního zatížení doktrínou TDL. • Na druhou stranu řada „hot-spots“ leží právě v TDL. • Situace ohledně ohrožení TDL je bohužel poměrně nejasná, různé druhy reagují na změny v TDL různě a stejně tak jsou antropogenní aktivity člověka v TDL velmi rozmanité (výběrové kácení stromů vs. plantáže palmy olejné v JV Asii, resp. pastviny pro chov dobytka v J. Americe). • Z hlediska ohrožení WD se jeví jako významnější stanoviště otevřena a bezlesá (stepi, savany, pampy, prérie, suché středomořské lesy a křoviny, opadavé prosýchavé tropické lesy apod.)