A Genomic Code for Nucleosome Positioning
250 likes | 879 Views
A Genomic Code for Nucleosome Positioning Authors: Segal E., Fondufe-Mittendorfe Y., Chen L., Thastrom A., Field Y., Moore I. K., Wang J.-P. Z., Widom J. Presented by Apostol Gramada DNA organization – Chromatin Taken from: http://sgi.bls.umkc.edu/waterborg/chromat/chroma09.html

A Genomic Code for Nucleosome Positioning
E N D
Presentation Transcript
A Genomic Code for Nucleosome Positioning Authors: Segal E., Fondufe-Mittendorfe Y., Chen L., Thastrom A., Field Y., Moore I. K., Wang J.-P. Z., Widom J. Presented by Apostol Gramada
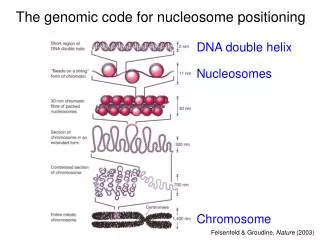
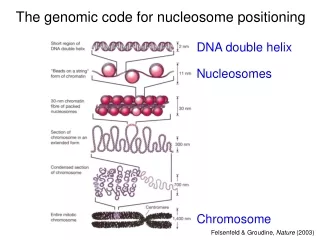
DNA organization – Chromatin Taken from: http://sgi.bls.umkc.edu/waterborg/chromat/chroma09.html
Nucleosome organization • An octamer of 8 histone chains, 2 of each of the following: H2A, H2B, H3, H4. • H3, H4 highly conserved in eukaryots. • 147 bp per nucleosome • DNA sharply bent and tightly wrapped in approx 1.7 turns around the histone core. • DNA bends discontinuously with the periodicity of the helical repeat. • Bending is facilitated by certain dinucleotides placed at the right positions.
Nucleosome organization • DNA in nucleosomes is far more sharply bent than in unstressed naked DNA => significant free energy cost needed for stability. • Particular DNA sequence could reduce this cost by • either having an inherent bendedness • being more easily bendable (more flexible). • The later seems to be more supported by evidence. • The ~10 bp periodicity of AA/TT, TA, GC seems to be an especially flexible sequence motif.
Nucleosome positioning • DNA sequences differ in their ability to bend sharply. This affects the DNA binding affinity of the histone octamer. • In vitro studies show a wide range of affinities with respect to sequence variability (approx 1000-fold). Some sequences highly preferred. • Is this mechanism used to control the access to specific binding sites? • The positions of the nucleosomes may have important inhibitory or facilitatory roles in regulating gene expression.
Nucleosome positioning – current views • Sequence preferences is over-ridden by nucleosome remodeling complexes which move them to new locations whenever needed. • Opposing view: the remodeling complexes only enable the nucleosomes to sample rapidly alternative positions and therefore compete efficiently with DNA binding proteins. They do not determine their destination however. Then, the genome would encode a nucleosome organization intrinsic to the DNA sequence alone, comprising sequences with both regions of low and high affinity for nucleosomes. • The high affinity regions will be occupied in vivo and the detailed distribution of nucleosome positions will significantly influence the chromosome functions genome-wide.
Validating a nucleosome-DNA interaction model • The data: 199 mono-nucleosome DNA sequences (142-152 bp) from yeast. • Used to construct a probabilistic model measuring the sequence preferences of yeast nucleosome: • Generate distribution functions at each site on the nucleosome for all dinucloetides, from the population of the 199 sequences. • A probability can then be assigned to each sequence of 147 bp. • Derive a thermodynamic model for predicting the nucleosome positions genome-wide from all legal configurations of nucleosomes (no overlap, at least 10 bp away).
Predicting nucleosome organization in genomic DNA sequence Resulting intrinsic nucleosome organizations: mutually exclusive organization dominate, a single organization dominate, none dominates => may reveal potential regulatory role of nucleosomes.
Predicted nucleosome organization reflects in vivo data • Orange Data in vivo. • 54% within 35 bp (only 39% by chance).
Predicted nucleosome organization reflects in vivo data • Comparison to three genome-wide measurements reveals: • significant correspondence between predicted and experimental nucleosome-depleted coding and intergenic regions: 68% of 57 depleted coding regions and 76% of 294 depleted intergenic regions. • strong correspondence with a higher resolution nucleosome map: 45% within 35bp distance (32% by chance).
Predicted nucleosome organization reflects in vivo data • Compared prediction of yeast model with one using only nucleosome-bound sequence from chicken
Global features of intrinsic nucleosome organization in yeast • From ~ 11 mil positions => ~15800 stable ncls. => cover 20% of genome. array? • Fig d shows the distribution of pairwise distances between stable ncls. => periodicity of ~ 177 bp extending over ~ six positions higher level chromatin organization?
Nucleosome organization varies by type of genomic region • Centromer function requires enhanced stability => max occupancy • Highly expressed Ribosomal RNA and transfer RNA => low predicted occupancy • Genes that very their expression levels (Ribosomal protein) in different conditions requires other mechanisms.
Nucleosomes facilitate their own remodeling • Analyzing ~1900 genes from a gene annotation database and various studies shows significance association with either high or low predicted occupancy • In particular, the chromatin remodeling complex RSC is associated with low occupancy => genomes facilitate their own remodeling
Low nucleosome occupancy encoded at functional binding sites • Stable ncls. over non-functional sites => decrease accessibility to transcription factors. • Tests showed: for 37% (out of 46) occupancy was lower over functional sites than for non-functional sites.
Low nucleosome occupancy encoded at transcription start sites
Conclusions • Nucleosome organization is encoded in eukaryotic genome • The limited predictive power (~50% of in vivo nucleosome organization) is explained by a too crude model yet: • a more accurate nucleosome-DNA interaction model • no account for favorable interactions and for steric hindrances implied by the 3D ncls structure • no account for competition with binding proteins.
Math Wc[S] = statistical weight of sequence S with nucleosome configuration c. “Legal” nucleosome configuration