Download
1 / 48
820 likes | 2.85k Views
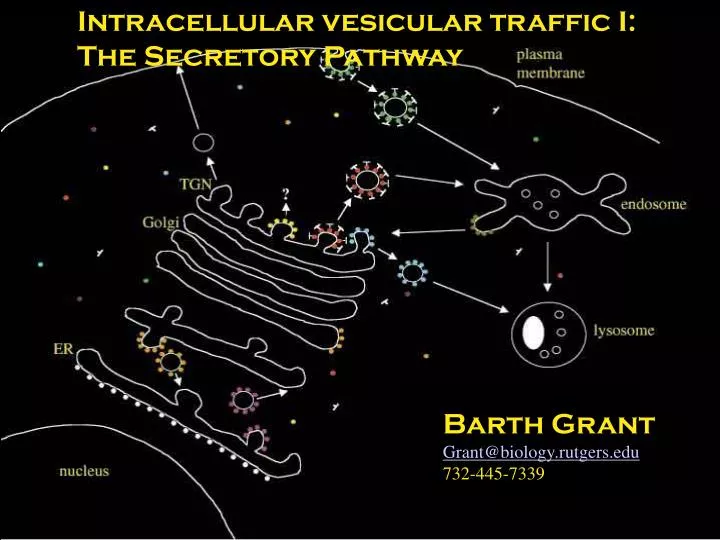
Intracellular vesicular traffic I: The Secretory Pathway. Barth Grant Grant@biology.rutgers.edu 732-445-7339. Compartments enclose a space called the lumen within a lipid bilayer. Vesicular Transport Proceeds by Budding and Fusion. Kinetics of Secretion. Overview of the secretory pathway.

E N D
Intracellular vesicular traffic I: The Secretory Pathway Barth Grant Grant@biology.rutgers.edu 732-445-7339
Compartments enclose a space called the lumen within a lipid bilayer. Vesicular Transport Proceeds by Budding and Fusion
Overview of the secretory pathway PM TGN All secretory and endocytic compartments are topologically equivalent to the extracellular space. Golgi Trans Medial Cis ERGIC ER
Green Fluorescent Protein(GFP) • The Beta-Can Structure • The structure of GFP has been determined at resolutions of 2.0 angstroms or better. (One angstrom is equal to one ten-billionth of a meter.) GFP is composed of a single chain of 238 residues, or amino acids, which form an 11-stranded beta-barrel wrapped around a central helix. The cylindrical barrel is 42 angstroms long and 24 angstroms in diameter, which protects both the central helix and the p-hydroxylbenzylideneimidazolidinone chromophore molecule that lays within). The chromophore is responsible for GFP's luminescence. • The chromophore absorbs blue light shoots off a proton and emits green light.
The Chromophore of GFP • GFP is unique among fluorescent proteins in that its fluorophore is not a seperately synthesized prostethic group but composed of modified amino acid residues within the polypeptide chain. • The green fluorochrome is formed by autocatalytic oxidation and condensation of residues ser65-tyr66-gly67 to form a conjugated ring system
Protein Folding and Complex Assembly Are Linked To Transport Protein Folding Chaperones like BIP/Hsp70 Prevent Unfolded Proteins and Unassembled Multimers such as antibodies from exiting the ER prematurely.
Vesicle Budding Utilizes Several Coat Protein Complexes • COPII mediates ER to Golgi transport • COPI mediates Golgi to ER transport • Clathrin mediates Golgi to Endosome transport, PM to Endosome transport, and perhaps other transport steps.
Vesicles Travel on Microtubules COPII-coated vesicles leave the ER, uncoat and begin to fuse with one-another to form vesicular-tubular clusters. These clusters associate with motor proteins that drag them along microtubules in an ATP dependent process. Meanwhile retrograde transport removes certain components, purifying and concentrating the secretory cargo further.
ERGIC to Golgi Transport - VSVG-GFP Microtubule Tracks and Motor Proteins Guide Transport Vesicles. Note the saltatory movement in straight lines.
Retrieval of ER Resident Proteins is Receptor-Mediated Escaped ER-resident proteins are retrieved from the Golgi by KDEL receptors that recognize specific retrieval signals in ER proteins.
ER-resident proteins often are retrieved from the cis-Golgi Lumenal proteins of the ER contain KDEL (Lys-Asp-Glu-Leu-COOH) Signals recognized by the KDEL receptor (ERD2). E.g. HSC70, PDI KDEL is necessary and sufficient. The KDEL-receptor cycles between the ER and Golgi via COPII and COPI-coated vesicles.
Early Secretory Pathway - Forward and Retrograde Traffic KDEL-receptors bind to KDEL-bearing proteins in the low pH environment of the Golgi and release that Cargo in the neutral pH of the ER. pH probably alters KDEL receptor conformation - regulating cargo binding and inclusion in COPI vesicles.
Post-translational Modification in the Golgi Note that specific modifications occur in specific subcompartments, because modifying enzyme localization is tightly controlled. The state of protein modification can identify how far a protein has proceeded in transport.
Mannose-6-Phosphate M6P modification is the key to sorting certain lysosomal enzymes during biosynthesis.
Mechanism of Mannose 6-phosphate addition GlcNAc phosphotransferase transfers N-acetylglucosamine phosphate to mannose on lysosomal Enzyme (e.g. Cathepsin D). A phosphodiesterase removes the GlcNAc, leaving the phosphorylated mannose. Phosphorylated mannose is a signal recognized by the Mannose-6-phosphate receptor.
M6P Sorting Pathway Late Endosome Lysosome
The mannose 6-phosphate (M6P) pathway Sorting of lumenal proteins can occur by binding transmembrane receptors. Lysosomal enzymes modified with M6P are bound by the lumenal domain of MP6R. MP6R-lysosomal enzyme complexes are recruited into clathrin/AP1 coated pits. Vesicles deliver the MP6R-lysosomal enzyme complexes to the late endosome. MP6R recycles to the golgi. Lysosomal enzymes are delivered to lysosomes.
Proproteins undergo proteolytic processing late in maturation
Two Models For Cis to Trans-Golgi Progression Tradional Model - Golgi is a static organelle. Secretory proteins move forward in small vesicles. Golgi resident proteins stay where they are. “Radical” Model - Golgi is a dynamic structure. It only exists as a steady-state representation of transport intermediates. Secreted molecules move ahead with a cisterna. Golgi resident proteins move backward to stay in the same relative position.
Randy Sheckman Analysis of yeast mutants defined the major steps in the secretory pathway Temperature sensitive lethal mutations define this pathway. Many novel proteins required for each of these steps were discovered in this genetic screen. Mutant phenotype reveals earliest requirement, not necessarily every requirement.
Class B Mutants Defined The COPII Coat Membrane and cargo molecules become concentrated in transport vesicles as they leave the ER. Membrane proteins interact with components of the COPII coat through exit signals on their cytosolic tails. Soluble proteins bind to certain concentrated membrane proteins known as cargo receptors.
Small GTPases Regulate Budding Sar1 is a small GTPase that acts to recruit coat proteins. Sar1-GDP is converted to Sar1-GTP by a transmembrane GEF. GTP binding triggers insertion of fatty acid tail of Sar1 into membrane. Sar1-GTP on the membrane recruits COPII proteins, resulting in membrane curvature. Pinching-off occurs to release the free vesicle. Uncoating occurs.
A cell-free system can be used to follow movement of proteins between Golgi cisternae Golgi from WT (uninfected) + Golgi from mutant cells (infected) + cytosol + ATP + 3H-N-acetylglucosamine. Measure 3H-modified viral protein as a measure of transport.
James Rothman Fractionation of “Cytosol” allowed biochemical purification of budding and fusion activities: COPI coat - budding, vesicle formation Snare Complex - membrane fusion NSF/Snap complex - snare recycling
COP I vesicles mediate retrograde transport within the Golgi and from the Golgi back to the ER 50 nanometer vesicles
Snares Provide Specificity To Fusion Vesicle SNARES (v-snares) and target membrane snares (t-snares) form cognate pairs that confer specificity on the fusion reaction. V-SNARES are incorporated into transport vesicles during budding. After fusion v-snares and t-snares remain tightly associated.
Specific fusion of intracellular vesicles involves a conserved set of fusion proteins SNARE proteins mediate both specificity of fusion and the fusion process itself. V-SNARE (vesicle), T-SNARE (target), and SNAP25 form a 4-helix bundle (coiled-coil). Specific Rab proteins are required for specific fusion events. Their exact role is unknown, but they probably function to recruit other effectors that participate in docking and fusion. Rab-GTP is the active form.
Snare Four-helix Bundle Neuronal snares of the synaptic plasma membrane have been particularly well characterized. The conserved snare complex is composed of a four-helix bundle. In this case one helix is contributed by the integral membrane v-snare synaptobrevin, one by the integral membrane t-snare syntaxin, and two by the peripheral membrane t-snare SNAP25.
NSF Unfolds SNARE Complexes, Allowing Them To Recycle NSF, a AAA-ATPase, associates with the snare complex via adaptor proteins and dissociates the complex in an ATP dependent manner. Without this reaction snares can be used only once!
Conformational changes in influenza HA protein trigger membrane fusion Virus binds to cell surface receptors modified with sialic acid. The “fusion peptide” is buried within the HA protein at neutral pH. (Spring-Loaded) The virus enters the endosomal pathway where the pH is lower. At pH 5 HA protein undergoes radical conformational change, extending the hyrophobic “fusion-peptide” into the target membrane, initiating fusion, releasing the viral DNA into the cytoplasm. The V-SNARE/T-SNARE/SNAP25 “snare-pin” resembles the HA “hairpin” . Binding of fusion peptide to HA2 disrupted. Globular domains dissociate. Loop segment forms a continuous helix. Fusion peptide inserts into endosomal membrane.
Model of membrane fusion directed by the viral HA protein HA insertion. Outer leaflet mixing. Bilayer fusion - fusion pore formation. Migration/widening of pore to complete fusion.
Membrane fusion as directed by SNAREs. Snare-pairing drives water from between opposing membranes. Lipids of the outer leaflets interact to form a stalk. The formerly inner leaflets meet to form a bilayer. This is called hemifusion. Rupture of the new bilayer creates the pores which expands to complete fusion.