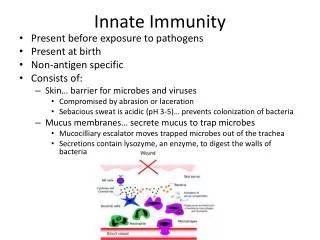
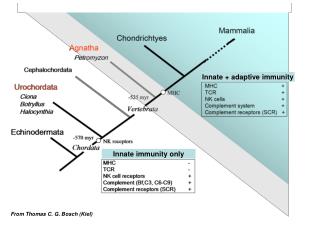
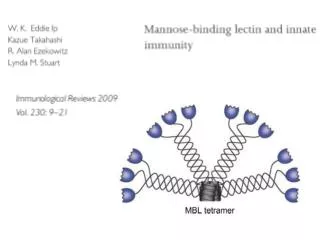
Download

1 / 6

E N D