Download
1 / 38
410 likes | 718 Views
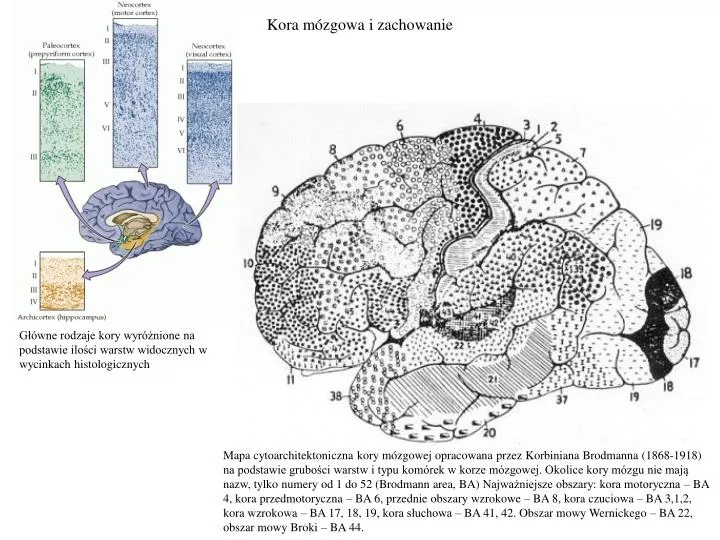
Kora mózgowa i zachowanie. Główne rodzaje kory wyróżnione na podstawie ilości warstw widocznych w wycinkach histologicznych.

E N D
Kora mózgowa i zachowanie Główne rodzaje kory wyróżnione na podstawie ilości warstw widocznych w wycinkach histologicznych Mapa cytoarchitektoniczna kory mózgowej opracowana przez Korbiniana Brodmanna (1868-1918) na podstawie grubości warstw i typu komórek w korze mózgowej. Okolice kory mózgu nie mają nazw, tylko numery od 1 do 52 (Brodmann area, BA)Najważniejsze obszary: kora motoryczna – BA 4, kora przedmotoryczna – BA 6, przednie obszary wzrokowe – BA 8, kora czuciowa – BA 3,1,2, kora wzrokowa – BA 17, 18, 19, kora słuchowa – BA 41, 42. Obszar mowy Wernickego – BA 22, obszar mowy Broki – BA 44.
Filogeneza kory mózgowej – wczesne poglądy • Wczesne (do ~1970) poglądy na wydarzenia w trakcie rozwoju kory mózgowej, od ryby do ssaka: • liniowy wzrost kresomózgowia (półkule mózgu+spoidła + jądra podstawne + węchomózgowie) • u ryby kora służyła głównie do odbioru (paleocortex) i analizy (archicortex) bodźców węchowych. • u płazów, a szczególnie u gadów, pojawiła się nowa kora (neocortex) • u gadów i ptaków inne bodźce sensoryczne dochodzą do kory poprzez wzgórze. • znaczne powiększenie kory nowej oraz ilości wejść ze wzgórza i zwojów podstawy u ssaków.
Filogeneza kory mózgowej – nowe obserwacje • Nowe obserwacje: • -Kresomózgowie nie rozwijało się stopniowo lecz skokowo (np. delfiny). • Projekcje dróg węchowych do kresomózgowia są właściwe (specyficzne) tylko dla pewnych obszarów kory. • Bodźce innych modalności sensorycznych, przechodzące przez wzgórze są obecne już u ryb • Podobnie, lecz niezależnie rozwinięte struktury nazwano ‘homoplastycznymi’ w odróżnieniu od odpowiednich struktur rozwijających się liniowo (tj. homologicznych).
Etapy rozwoju ludzkiego mózgu: • 4 tygodnie: • - widoczny podział cewki nerwowej (neural tube)na różne obszary (prosencephalon, encephalon, mesencephalon, rhomboencephalon) • pojawia się pęcherzyk wzrokowy (optic vesicle, OV) • 5 tygodni: • wyróżniają się nowe obszary: telencephalon i diencephalon, metencephalon i myelencephalon • Powstaje zalążek podwzgórza • 7 tygodni: • -powstaje wzgórze łączące telencephalon z pozostałymi częściami układu nerwowego. • -powstaje wyspa (medulla), most (pons) i móżdżek (cerebellum) • ------------------------------------------------------------------zmiany typowe dla ssaków: • 2 miesiące: • wzrost telencephalonu • 3 miesiące: • -telencephalon przykrywa część ogonową pnia mózgu • widoczny początek podziału na płaty (lobes) • 4 - 9 miesięcy: • wzrost kory nowej wraz z pofałdowaniem • zupełne przykrycie rdzenia kręgowego Ontogeneza kory mózgowej
Początki układu nerwowego We wczesnym zarodku zaczynają się wyróżniać 3 warstwy: zewnętrzna (ektoderma), wewnętrzna (endoderma) i środkowa (mezoderma). Układ nerwowy rozpoczyna się we wczesnym stadium rozwoju embrionalnego w części środkowogrzbietowej ektodermy - tzw. płytce nerwowej.
Początki układu nerwowego • Położenie płytki nerwowej w stosunku do nieneuronalnej ektodermy, mesodermy i endodermy. • Komórki grzebienia nerwowego (neural groove) leżące po obu stronach płytki nerwowej rozpoczynają różnicowanie się obwodowego układu nerwowego. • Rynienka zagłębia się coraz bardziej • Wierzchołki rynienki stykają się ze sobą tworząc cewę nerwową. Komórki wyściełające cewę nerwową dzielą się i różnicują w kierunku neuronów i komórek glejowych.
Neurogeneza – powstawanie neuronów Komórki wyściełające cewę nerwową przechodzą powtarzalny cykl migracji związanych z podziałem komórki (mitozą). Cykl rozpoczyna się od niezróżnicowanej komórki zawierającej diploidalne DNA (zawierająca po dwa chromosomy danego typu ) (faza G1). Jądro komórki przemieszcza się na zewnątrz i rozpoczyna replikacje DNA (faza S). Po podwojeniu zawartości DNA w jądrze (dolny rysunek), przemieszcza się ono z powrotem do wewnątrz (faza G2) i rozpoczyna mitozę w pobliżu ścianki (faza M). W wyniku mitozy powstają dwie komórki z diploidalną zawartością DNA. Powstała komórka i) może stać się neuronem i rozpocząć migrację w kierunku swojego miejsca docelowego, lub ii) staje się komórką niezróżnicowaną i zaczyna od nowa powtarzalny cykl podziału.
Migracje neuronów Migrujący neuron. Z: http://www.rockefeller.edu/labheads/hatten/mechanism.html Radialne komórki glejowe dostarczają wytycznych dla migrujących neuronów W płytce korowej (cortical plate, CP) neurony mijają warstwy głębokie i dążą do warstwy I.
Dojrzewanie neuronów Dojrzewanie neuronów trwa znaczną część życia. Podczas rozwoju neuronu od fazy embrionu do dorosłego człowieka, neurony rosną, oraz wykształcają drzewa dendrytyczne i kolaterale aksonu. Wczesne dzieciństwo jest fazą dynamicznego dojrzewania komórek w korze.
Dojrzewanie synaps Przebieg czasowy rozwoju synaps w różnych obszarach kory mózgowej małpy. Na rys. F pokazano lokalizacje badanych obszarów. Powierzchniowa gęstości synaps w neuropilu (tkanka korowa bez ciał komórek i bez naczyń krwionośnych) była badana poprzez liczenie synaps na 100 mikrografiach z mikroskopu elektronowego. Poziomy szary pas wskazuje wartość średnią u dorosłego osobnika. Czas (w skali logarytmicznej) przebiegał od zapłodnienia do wieku 20 lat. W badaniach zidentyfikowano 500 000 synaps w 25 000 mikorgrafiach u 22 małp. Schemat rozwoju synaps jest taki sam we wszystkich badanych obszarach kory. U płodu następuje stały wzrost gęstości synaps, osiągający poziom dojrzałego organizmu w chwili urodzin. Po narodzinach gęstość rośnie jeszcze przez 2-4 miesiące, a następnie opada (wpierw szybko, następnie wolno) i osiąga poziomu dorosły w wieku 2-4 lata. Wyniki wskazują na ‘nadprodukcję i eliminację’. Konkurencja i walka o przetrwanie w świecie zewnętrznym odzwierciedla się w walce o przetrwanie synaps w wewnętrznym świecie neuronów. ‘Przeżywają’ obwody najbardziej skuteczne w przetrwaniu całego organizmu.
Poziomy organizacji kory Poziomy organizacji układu nerwowego i kory mózgowej. Poziom cząsteczek i jonów oraz kanałów i receptorów w korze odpowiada tym samym poziomom w innych strukturach mózgu.
Poziomy organizacji kory – synapsy i kolce dendrytyczne Rozwój synaptyczny jest w dużej mierze związany z rozwojem kolców dendrytycznych. Każdy kolec jest ‘siedzibą’ synapsy; kolce odgrywają role w ‘nadprodukcji i eliminacji’. Synapsy w korze są bardzo czułe na sygnały środowiska. Np. pozbawienie sygnałów wzrokowych prowadzi do redukcji ilości kolców (w ciele kolankowatym bocznym - część B na rysunku. 1 – warunki normalne, 2 –wychowanie w ciemności), wychowanie w stymulującym otoczeniu powoduje wzrost grubości kory i większe połączenia synaptyczne.
Poziomy organizacji kory – jednostki kolców dendrytycznych Oddziaływanie pomiędzy kolcami może generować operacje logiczne w dendrytach komórek kory. A. Segment dendrytyczny neuronu kory z czterema wejściami synaptycznymi. B. Model kompartmentowy segementu z rys. A. Kompartmenty pasywne pokazane są na biało, kompartmenty aktywne są zacieniowane, a C i D przedstawia ich obwody zastępcze. E. Symulacja odpowiedzi układu w wyniku pobudzenia komp. 1 i 2. Jednoczesne pobudzenie 1 i 2 prowadzi do powstania impulsów w komp. 1 i 2 oraz, poprzez pasywną propagację, w komp. 3 i 4. Układ funkcjonuje jak bramka logiczna AND. F. Symulacja odpowiedzi układu w wyniku pobudzenia komp. 1 i hamowania komp. 7. Hamowanie komp. 7 blokuje powstawanie impulsu w komp. 2 i 3 lecz nie w komp. 1. Układ funkcjonuje jak bramka logiczna NOT-AND
Poziomy organizacji kory – neurony korowe W korze znajduje się wiele rodzajów komórek. Występuje tu zasadniczy podział na komórki piramidalne, tworzące wyjścia i niepiramidalne, głównie interneurony hamujące. Komórki w różnych warstwach kory różnią się rozmiarem i własnościami. W płytkich warstwach kory występują komórki typu RS (regular spiking), w głębszych warstwach występują zarówno RS jak i IB (intrinsically bursting). Interneurony we wszystkich warstwach są typu FS (fast spiking). Funkcjonalne znaczenie różnych wzorców odpalania jest wciąż badane. Mircea Steriade, Neocortical Cell Classes Are Flexible Entities. NATURE REVIEWS |NEUROSCIENCE, VOL. 5, pp. 121-134, 2004.
Poziomy organizacji kory – obwody lokalne Podobnie jak w innych obszarach mózgu, obwody synaptyczne w korze zbudowane są z elementów triady synaptycznej: włókna wejściowe, neurony wyjściowe i neurony wewnętrzne (interneurony). A. Podstawowy obwód korowy: 1. Dostaje wejścia pobudzające na dendryty dystalne neuronów piramidalnych. 2. Kolaterale axonalne są pobudzające (reexcitatory, RE). 3. Interneurony dostają wejścia i zapewniają hamowanie do przodu (feedforward inhibition, FI) i/lub otrzymują wejścia z kolaterali i zapewniają hamowanie boczne (lateral inhibition, LI). W prostej korze (agranular neocortex, B.) np. kora motoryczna i asocjacyjna, komórki dostają wejścia z innych obszarów kory. Występują połączenia typu RE, FI, LI. W korze ziarnistej (granular neocortex, C.) np. kora wzrokowa, wejścia aferentne ze wzgórza dochodzą do komórek gwiaździstych (stellate cells), definiując nową warstwę (IV). Komórki gwiaździste tworzą stację przekaźnikową w korze, w której odbywa się wstępne obrabianie informacji, zanim dotrze ona do komórek piramidowych.
Poziomy organizacji kory – moduły Obwody lokalne w korze nie są zorganizowane w sposób rozmyty lecz w skupiska tzw. moduły. Przykłady modularyzacji kory podane są w tabeli.
Poziomy organizacji kory – moduły w korze wzrokowej Moduły nie są wpisane statycznie lecz tworzą się pod wpływem doświadczenia. A. U małpy projekcje oczne z ciała kolankowatego bocznego w embrionie są początkowo rozległe i nakładające się na siebie w korze wzrokowej. Na 2 tygodnie przed urodzeniem (E144) następuje segregacja wejść z jednego i drugiego oka i powstanie kolumn dominacji ocznej, w pełni ukształtowanych w 3-6 tygodni po urodzeniu. B. Jeśli jedno oko będzie zamknięte od urodzenia, kolumny dominacji ocznej drugiego oka ulegną powiększeniu. Deprywacja jednooczna następowała w wieku: a) 2 tygodnie, b) 5.5 tygodnia, c) 10 tygodni, d) w dorosłości. Zmiany w korze wzrokowej następują gdy deprywacja zaczyna się w trakcie kształtowania się kolumn (przypadek a i b).
Poziomy organizacji kory – obszary i płaty korowe Kora ssaków składa się z czterech płatów: czołowy (frontal), ciemieniowy (parietal), skroniowy (temporal) i potyliczny (occipital). Płat potyliczny dostaje wejścia wzrokowe, płat ciemieniowy – wejścia czuciowe, płat skroniowy – wejścia słuchowe, w płacie czołowym ma początki wiele dróg ruchowych. Obszary płatów nie zaangażowane bezpośrednio w drogi zmysłowe lub motoryczne nazywane są obszarami asocjacyjnymi. Obszary te uległy największemu powiększeniu u człowieka co czyni je jednym z atrybutów człowieka.
Poziomy organizacji kory – układy rozproszone – przetwarzanie informacji wzrokowej Większość zachowań kontrolowanych przez korę zależy od oddziaływania obszarów należących do różnych płatów korowych. Innymi słowy, funkcje kory są zorganizowane w systemy rozproszone. Przykładem może być analizowanie informacji wzrokowej angażujące obszary w płacie potylicznym, ciemieniowym i skroniowym. B.W niższej części płata skroniowego analizowane są własności obiektów. C.W tylnej części płata ciemieniowego analizowane są relacje przestrzenne między obiektami. Droga grzbietowa (dorsal) jest wielosensoryczna i łączy informacje wzrokową z informacją czuciową ręki.
Wewnętrzne reprezentacje i płaty czołowe A. W płatach czołowych następuje tworzenie wewnętrznej reprezentacji świata zewnętrznego na podstawie informacji wzrokowej. Małpy z uszkodzoną korą czołową (i noworodki do 1 roku życia) nie potrafią wykonać zadania polegającego na utrzymaniu obiektu w wewnętrznej reprezentacji, nie znajdującego się w polu widzenia (delayed response test). B. Badania neuroanatomiczne pokazały silne wzajemne połączenia pomiędzy obszarami asocjacyjnymi w korze przedczołowej i ciemieniowej. Występują też drogi równoległe biorące udział w analizowaniu różnego typu informacji.
Płaty czołowe, pamięć robocza i choroby psychiczne „Working memory confers the ability to guide behavior by internal representations of the outside world rather than by immediate stimulation and thus to base behavior on ideas and thoughts.” P. Goldman-Rakic, 1991. Prefrontal cortical disfunction in schizophrenia: the relevance of working memory. In Psychopathology and the Brain. New York, Raven Press. Schizofrenia może polegać na załamaniu mechanizmów dzięki którym reprezentacja wewnętrzna świata (wiedza) rządzi zachowaniem. Obserwacje potwierdzające hipotezę: 1.Schizofrenicy łatwo się rozpraszają pod wpływem zewnętrznych bodźców 2. Schizofrenicy słabo wypadają w teście delayed response 3. Pacjenci ze schizofrenią mają zmniejszony przepływ krwi w płatach czołowych.
Półkule mózgowe – lateralizacja i dominacja Pierwszy dowód na asymetrię funkcjonalną (lateralizacja) półkul podał Paul Broca w 1863. Aparat mowy znajduje się w płacie czołowym lewej półkuli (dominacja lewej półkuli w mowie). Oryginalny rysunek Wernickego (1874) pokazujący koncepcje obwodów mowy. Dźwięk docierający do uszu, zamienia się na impulsy nerwowe i dociera drogami słuchowymi (a) do mózgu, gdzie ‘obrazy’ dźwięku są przechowywane w polu Wernickego (a’). Obrazy neuronalne przesyłane są do pola Broki (b), które aktywuje wychodzące projekcje motoryczne kontrolujące aparat mowy.
Dominacja Schematyczny rysunek mózgu z zaznaczonymi specjalizacjami obu półkul. Pomiędzy półkulami widoczne przecięte spoidło wielkie. Lewa półkula dominuje w: mowie, języku, złożonych ruchach zamierzonych, czytaniu, pisaniu i arytmetyce. Prawa półkula dominuje w: funkcjach nielingwistycznych, takich jak rozpoznawanie wzorców wzrokowych, słuchowych, czuciowych, orientacji przestrzennej, rozpoznawanie kształtu, orientacji kierunku w przestrzeni oraz intuicji. Ogólny podział: półkula lewa – naukowa, półkula prawa - artystyczna
Lateralizacja Oprócz asymetrii funkcjonalnej odkryto (w 1968 r.) asymetrię anatomiczną związaną z obszarami mowy w mózgu. Obszar zwany ‘Planum Temporale’, zawierający ośrodek Wernickego jest znacząco większy z lewej strony.
Wypowiadanie wyrazów Ośrodki mózgu zaangażowane w wypowiadanie nazw widzianych obiektów. Informacja wzrokowa dochodzi do pola 17 i jest przetwarzana w polach 18 i 19. Stamtąd obraz percepcyjny jest przekazywany do ciemieniowego obszaru mowy zawierającego pole 39 i klasyczny obszar Wernickego. Pole 39 przekazuje wzrokową reprezentacje obiektu do pola 22, gdzie powstaje reprezentacja słuchowa. Z pola 22, informacja dostaje się do pola Broki, gdzie uruchamiane są programy motoryczne mowy. Programy te kontrolują czasowo-przestrzenną koordynacje mięśni mowy i są odpowiedzialne za wypowiadane słowo.
PET – technika obrazowania, w której rejestruje się promieniowanie powstające w podczas anihilacji pozytonów. Źródłem pozytonów jest substancja promieniotwórcza podana badanemu np. deoxyglukoza znakowana izotopem fluoru (F-18). W miejscach podwyższonego metabolizmu, następuje spalanie cukrów – tam też gromadzi się deoxyglukoza, będąca ‘źródłem’ pozytonów. Pozytony, przebywają drogę ~mm i zderzają się z elektronami. W wyniku anihilacji powstają dwa fotony poruszające się w przeciwnych kierunkach.Są one rejestrowane jednocześnie przez dwa z wielu detektorów ustawionych wokół (najczęściej w postaci pierścienia) badanego organu, w wyniku czego można określić dokładne miejsce powstania pozytonów. Informacje te rejestrowane w postaci cyfrowej pozwalają na konstrukcję obrazów będących przekrojami ciała (mózgu) pacjenta. Pozytonowa Tomografia Emisyjna (Positron Emission Tomography, PET Badanie aktywności mózgu wykonuje się w warunkach kontrolnych i podczas wykonywania zadania. Różnica obrazów pokazuje aktywność związaną z zadaniem. Uśrednianie wielu obrazach prowadzi do redukcji szumu.
Myślący mózg – analiza informacji słownej Badanie aktywności mózgu metodą PET podczas wykonywania trzech różnych zadań o rosnącej złożoności. Dla każdego zadania obraz aktywności prostszego zadania był odejmowany od obrazu zadania bardziej skomplikowanego by wyróżnić miejsca w mózgu odpowiedzialne jedynie za zadanie skomplikowane. Zadanie 1: pasywny odbiór bodźca sensorycznego – słowa prezentowanego A – wzrokowo, B – słuchowo, w porównaniu z fiksacja wzrokową. Zadanie 2: wypowiadanie prezentowanego słowa C), w porównaniu z pasywnym odbiorem. Zadanie 3: wypowiadanie słowa skojarzonego ze słowem prezentowanym (D), w porównaniu z wypowiadaniem prezentowanego słowa. Z: Petersen SE, Fox PT, Posner MI, Mintun M, Raichle ME.Positron emission tomographic studies of the cortical anatomy of single-word processing.Nature. 1988 18;331(6157):585-9.
Jak kodowana jest informacja w mózgu za pomocą potencjałów czynnościowych? Kod Mózgu Binarnie Częstość – ilość potencjałów czynnościowych na jednostkę czasu (rate coding): narządy zmysłów, mięśnie
Kod Mózgu Kod czasowy – zależność czasowa między impulsami
Kod Mózgu Kodowanie populacyjne – wzorzec aktywności w populacji neuronów. Np. kodowanie kierunku ruch, orientacji przestrzennej bodźca, smaku, koloru
Gamma coding Singerand von der Malsburg: gamma oscillation is a network phenomenonin which a group of cells fire together synchronously (synchronous is defined as firing in samegamma cycle; different groups fire on differentgamma cycles.) Cognitiverole: cells representing groups of features thatwere part of the same object would fire synchronously (‘‘binding’’). Different assemblies firing in differentgamma cycles could represent different entities (A, B, C..) that are simultaneously encoded in the brain. Singer W. 2000. Response synchronization: a universal coding strategy forthe definition of relations. In: Gazzaniga MS, editor. The new cognitiveneurosciences. 2nd ed. Cambridge, MA: MIT Press. p 325–338.
Theta coding – phase precession in hippocampal cells Hippocampal place cells code for spatial location. Code: phase of a spike with respect to theta cycle. 180o As the rat traverses the place fieldthe phase of each spike relative to thepeaks of theta in the field potential changes systematically. 0o 360o Mehta MR, Lee AK, Wilson MA. 2002. Role of experience and oscillationsin transforming a rate code into a temporal code. Nature417:741–746. O’Keefe J, Recce ML. Phase relationship between hippocampalplace units and the EEG theta rhythm. Hippocampus1993;3:317–30.
Theta coding – phase precession in hippocampal cells Prediction: that two cells coding for slightly different positionsshould show a cross-correlation with a peak offset equal to a significantfraction of theta. Confirmed! Cross-correlation between 4 place cells (pairwise1–2,1–3, and 1–4).Different cells havesystematic theta phase difference. The peak cross-correlation (lower panels) has a temporaloffset in the 40–80 ms range. Skaggs WE, McNaughton BL, Wilson MA, Barnes CA. 1996. Thetaphase precession in hippocampal neuronal populations and thecompression of temporal sequences. Hippocampus 6:149–172.
Theta/gamma coding – working memory model Each theta cycle is subdividedinto 7 gamma subcycles (A-G). Within a particular gamma cycle (e.g., A), certain cells will be active,forming a spatial code for an item. Seven items are sequentiallyrepresented in the gamma subcycles. The entire pattern group of 7 patternsmay repeat on the next theta cycle if this is a working memorynetwork. Decoding: coincidence detection by downstream cells Lisman J, The Theta/Gamma Discrete Phase Code Occuring Duringthe Hippocampal Phase Precession May be a MoreGeneral Brain Coding Scheme. HIPPOCAMPUS 15:913–922 (2005)
Gamma vs. Theta/gamma coding A B A B Order is relative gamma theta but with theta theta + gamma becomes absolute Order is BA! Singer’s coding scheme depends only on gamma. What it is that the presenceof theta provides? Suppose two assemblies (A,B) fire in sequentialgamma cycles. What is the orderAB or BA? Theta provides absolute phase reference and can conveyinformation about position in an ordered set Theta is a brain clock fora neural code designed for handling ordered units of information.
Theta/gamma coding in humans -analysis of EEG, MEG and intracranial EEG recordings -experiments motivated by animal studies -behavioral (verbal, visual and spatial cognition) correlates of gamma and theta oscillations were found often depending on brain region and poststimulus latency Increase of theta power during long maze navigation at selected brain sites: Construction of an object representation induces gamma burst at 280 ms triangle no triangle Tallon-Baudry C, Bertrand O, Delpuech C, Pernier J.Stimulus specificity of phase-locked and non-phase-locked 40 Hz visual responses in human.J Neurosci. 1996;16(13):4240-9. Kahana MJ, Sekuler R, Caplan JB, Kirschen M, Madsen JR: Humantheta oscillations exhibit task dependence during virtual mazenavigation.Nature 1999, 399:781-784.