Download

1 / 37
370 likes | 479 Views
Control of Transcription Initiation General References Chapter 16 of Molecular Biology of the Gene 6 th Edition (2008) by Watson, JD, Baker, TA, Bell, SP, Gann, A, Levine, M, Losick, R. 547-587.

E N D
Control of Transcription Initiation General References Chapter 16 of Molecular Biology of the Gene 6th Edition (2008) by Watson, JD, Baker, TA, Bell, SP, Gann, A, Levine, M, Losick, R. 547-587. Ptashne, M. and Gann, A. (2002) Genes and Signals. Cold Spring Harbor Laboratory Press, Cold Spring Harbor. Luscombe, N.M., Austin, S.E., Berman, H.M., Thornton, J.M. (2000) An overview of the structures of protein-DNA complexes. Genome Biology 1(1): reviews001.1-001.37 Examples of Control Mechanisms Alternative Sigma Factors Sorenson, MK, Ray, SS, Darst, SA (2004) Crystal structure of the flagellar sigma/anti-sigma complex 28 /FlgM reveals an intact sigma factor in an inactive conformation. Molecular Cell14:127-138. Gruber, TM, Gross, CA (2003) Multiple sigma subunits and the partitioning of bacterial transcription space. Annu. Rev. Microbiol57:441-66 Increasing the Initial Binding of RNA Polymerase Holoenzyme to DNA Lawson CL, Swigon D, Murakami KS, Darst SA, Berman HM, Ebright RH. (2004) Catabolite activator protein: DNA binding and transcription activation. Curr Opin Struct Biol. 14:10-20. Increasing the Rate of Isomerization of RNA Polymerase *Dove, S.L., Huang, F.W., and Hochschild, A. (2000) Mechanism for a transcriptional activator that works at the isomerization step. Proc Natl Acad Sci USA97: 13215-13220. Jain, D. Nickels, B.E., Sun, L., Hochschild, A., and Darst, S.A. (2004) Structure of a ternary transcription activation complex. Mol Cell 13: 45-53. Hawley and McClure (1982) Mechanism of Activation of Transcription from the l PRM promoter. JMB 157: 493-525
DNA looping **Oehler, S., Eismann, E.R., Kramer, H. and Mueller-Hill, B. (1990) The three operators of the lac operon cooperate in repression. EMBO 9:973-979. Vilar, J.M.G. and Leibler, S. (2003) DNA looping and physical constraints on transcription regulation. J Mol Biol 331:981-989. Dodd, I.B., Shearwin, K.E., Perkins, A.J., Burr, T., Hochschild, A., and Egan, J.B. (2004) Cooperativity in long-range gene regulation by the cI repressor. Genes Dev. 18:344-354. *Choi, PJ, Cai,L, Frieda K and X. Sunney Xie (2008) A Stochastic Single-Molecule Event Triggers Phenotype Switching of a Bacterial Cell Science 2008: 442-446. [DOI:10.1126/science.1161427] Chromosome conformation capture (CCC) de Wit, E. and de Laat, W. (2012) A decade of 3C technologies: insights into nuclear organization. Genes Dev. 26: 11-24. Mediator and Other Components *Kornberg, R.D. (2005) Mediator and the mechanism of transcriptional activation. Trends in Biochemical Sciences 30:235-239. Fan, X, Chou, DM, & Struhl, K. (2006). Activator-specific recruitment of Mediator in vivo. Nature Structural & Molecular Biology, 13(2), 117-20. Sikorski TW and Buratowski. (2009). The basal initiation machinery: Beyond the general transcription factors. Current Opinion in Cell Biology. 21 344-351. What do activators do? Cosma, MP, Tanaka, T, & Nasmyth, K. (1999). Ordered recruitment of transcription and chromatin remodeling factors to a cell cycle- and developmentally regulated promoter. Cell, 97(3), 299-311. Bryant, GO, & Ptashne, M. (2003). Independent recruitment in vivo by Gal4 of two complexes required for transcription. Molecular Cell, 11(5), 1301-9. Bhaumik, S.R., Raha, T. Aiello, D.P., and Green, M.R. (2004) In vivo target of a transcriptional activator revealed by fluorescence resonance energy transfer. Genes Dev 18: 333-343. Vakoc, CR, Letting, DL, Gheldof, ... Blobel, GA (2005) Proximity among Distant Regulatory Elements at the B–Globin Locus Requires GATA-1 and FOG-1. Molecular Cell 17:453-462 Fishburn, J., Mohibullah, N. and Hahn, S. (2005) Function of a eukaryotic transcription activator during the transcription cycle. Molecular Cell 18:369-378. Bulger M and Groudine M. Functional and Mechanistic Diversity of Distal Transcription Enhancers (2011). Cell 144:327-39
Important Points • 1. Every step in transcription initiation can be regulated to increase or decrease the number of successful initiations per time. • 2. In E. coli, transcription initiation is controlled primarily by alternative factors and by a large variety of other sequence-specific DNA-binding proteins. • 3. G=RTlnKD. This means that a net increase of 1.4 kcal/mole (the approximate contribution of an additional hydrogen bond) increases binding affinity by 10-fold. Many examples of transcription activation in bacteria take advantage of such weak interactions. • 4. To activate transcription at a given promoter by increasing KB, the concentration of RNA polymerase in the cell and its affinity for the promoter must be in the range so an increase in KB makes a difference. Likewise, to activate transcription by increasing kf, the rate of isomerization must be slow enough so the increase makes a substantial difference. • 5. DNA looping allows proteins bound to distant sites on DNA to interact. • 6. Transcription initiation at Pol II promoters on naked DNA templates in vitro requires the general transcription factors in addition to RNA polymerase II. • 7.. In vivo, transcription initiation also requires activators – proteins that bind directly to enhancers – as well as Mediator and enzymes that modify chromatin structure. • 8. At a typical eukaryotic promoter, activators guide the assembly of Mediator, the general transcription factors, RNA polymerase and chromatin-modifying enzymes, often through weak, relatively non-specific interactions. There appears to be no set order of assembly from one promoter to the next. Moreover, different promoters have different requirements for these components.
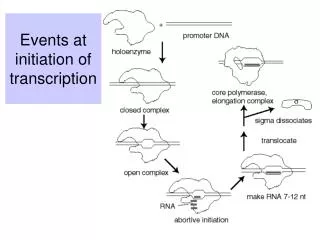
NTPs KB Kf Elongating Complex Abortive Initiation R+P RPc RPo initial binding “isomerization” Every step of transcription can be regulated
Gene regulation in E. coli: The Broad Perspective • 4400 genes • 300-350 sequence-specific DNA-binding proteins • 7 factors Alternative s are major control mechanism in bacteria
Regulation by repressors and activators (alter reactivity of s70-holoenzyme)
In vivo parameters for Sequence-Specific DNA binding proteins KD ≈ 10-6 - 10-10M in vivo In E. coli 1 copy/cell ≈ 10-9 M If KD = 10-9M and things are simple: 10 copies/cell 90% occupied 100 copies/cell 99% occupied Common families of DNA binding proteins
Negative control: repressors (e.g. , Lac ); prevent RNAP binding R -35 -10 Positive control: activators ( e.g. CAP); facilitate RNAP binding with favorable protein-protein contact * RNAP holo A Favorable contact -35 -10 I. Regulating transcription initiation at KB(initial binding) step
Lac ~ 1980 -35 -10 O3 O1 O2 Lac operator -90 -10 +400 -35 Lac 2000 Oehler, 2000 O2 1/10 affinity of O1 O3 1/300 affinity of O1 Lac repressor and DNA looping What is the function of these weak operators?
The weak operators significantly enhance represssion Oehler, 2000
Om Better! Oa M M A mutant Lac repressor that cannot form tetramers is not helped by a weak site Om Through DNA looping, Lac repressor binding to a “strong” operator (Om) can be helped by binding to a “weak” operator (OA) OK
Theoretical consideration of effects of looping (2 operators) Representative states of the binding of the repressor to one operator (top panel) or to two operators (bottom panel). Om (main operator) binds repressor more tightly than Oa (auxiliary operator). Transcription takes place only in the states (i) and (iii), when Om is not occupied. The arrows indicate the possible transitions between states. Note that with one operator, a single unbinding event is enough for the repressor to completely leave the neighborhood of the main operator. With two operators, the repressor can escape from the neighborhood of the main operator only if it unbinds sequentially both operators. From: Vilar, J.M.G. and Leibler, S. (2003) DNA looping and physical constraints on transcription regulation. J Mol Biol 331:981-989 .
Positive control: activators ( e.g. CAP); facilitate RNAP binding with favorable protein-protein contact * RNAP holo A Favorable contact -35 -10 I. Regulating transcription initiation at KB(initial binding) step ∆ G = RT lnKD; if * nets 1.4 kcal/mol, KB goes up 10-fold
RNAP a) If initial occupancy of promoter is low 1% occupied * A RNAP 10% occupied RNAP b) If initial occupancy of promoter is high 99% occupied * RNAP A 99.9% occupied Activating by increasing KB is effective only if initial promoter occupancy is low If favorable contact nets 1.4Kcal/mole (KB goes up 10X) then: Transcription rate increases 10-fold Little or no effect on transcription rate
cAMP high glucose How is CAP activated? Inactive CAP Active CAP Regulates >100 genes positively or negatively CAP at lac operon CAP increases transcription ~40-fold; KB ; no effect on kf A case study of activation at KB: CAP at the lac operon:
M M 1. Isolate “positive control” (pc) mutations in CAP. These mutant proteins bind DNA normally but do not activate transcription 2. “Label transfer” (in vitro) from activator labeled near putative “pc” site to RNAP S-S-X* RNAP RNAP Activate X*; reduce S-S; X* is transferred to nearest site; determine location by protein cleavage studies; X* transferred to -CTD -35 -10 M 3. Isolate CAP-non-responsive mutations in -CTD -35 -10 Strategies to identify point of contact between CAP and RNAP
Signal transduction cascade Signal Transcription factor Genes controlled by transcription factor
Common network motifs in transcriptional circuits Using a feed-forward loop to measure signal duration
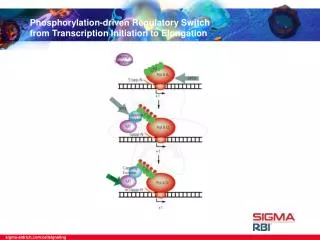
Transcription Initiation by RNA Pol II The stepwise assembly of the Pol II preinitiation complex is shown here. Once assembled at the promoter, Pol II leaves the preinitiation complex upon addition of the nucleotide precursors required for RNA synthesis and after phosphorylation of serine resides within the enzyme’s “tail”.
VP 16 GAL4 crude lysate 10 units 4 years mediator 50 units The GTFs are not sufficient to mediate activation: Discovery and isolation of Mediator from Yeast GTFs and RNA Pol II Tx 1 unit 1 unit
Control: ts subunit of Pol II Rpb1 high T Compare levels of all mRNAs using microarrays (37˚C for 45 min) Rpb1ts Subunit of Mediator Same basic pattern Srb4ts Subunit of TF II H Same basic pattern Kin28ts Is Mediator Required for Transcription of all Pol II -transcribed genes? mRNAs decrease over time according to their half-lives. A few mRNAs remain at some level (stable mRNAs).
Mediator is very large and has diverse roles PIC model from EM-study of polII (brown)-TFIIF (light blue) and X-ray structure of TBP (white) -IIB (yellow)--DNA; Arrow indicates direction of trx Model for PIC-mediator was produced by superimposing an EM structure of Mediator-PolII on the PIC in A; head, middle and tail regions shown The Head region interacts with PolII-TFIIF complex; the mutants with general effects on Trx are located in this region; the tail region interacts with activators; mutants have more specific effects on transcription
SAGA is another important complex with multiple roles in transcription, including being a coactivator The core of SAGA, containing the Taf substructure (Yellow), is surrounded by three domains responsible for distinct functions: activator binding (Tra-1), histone acetylation Gcn5), and TBP regulation (Spt3). This structural organization illustrates an underlying principle of modularity that may be extended to our understanding of other multifunctional transcription complexes.
Assembly of PIC in presence of mediator, activators and chromatin remodelers
Many Paths to the PIC Buratowski, 2009 The factors and assembly pathways used to form transcriptionally competent preinitiation complexes can be promoter dependent. (1) TBP assembling onto promoter regions via TFIID leads to recruitment of the other basal initiation factors, as outlined in the stepwise assembly pathway. In S. cerevisiae, this pathway is most often utilized at TATA-less genes. At some mammalian promoters, histone H3K4 trimethylation helps to recruit the TFIID complex. (2) Mediator bridges interactions between activators and the basal initiation machinery, and can stimulate basal transcription as well. At some promoters Mediator can recruit TFIIH and TFIIE independently of RNApoII. (3) TBP can also be brought to promoters by the SAGA complex. In S. cerevisiae, this pathway is most utilized at TATA containing promoters. The Mot1 and NC2 complexes can repress this pathway by actively removing TBP from the TATA element. (4) Mot1 and NC2 can also have a positive role in transcription by removing non-productive TBP complexes from DNA, thereby allowing functional PICs to form.
Genomic Level Snapshots of transcription factor binding sites ChIP ChIP-ChIP ChIP-seq ChIP-exo Bioinformatic approaches Phylogenetic footprinting
~ 30-100 bp ~ 100s bp Could be 50kB or more Regulatory sequences expand in number and complexity with increased complexity of the organism
3C is a variant of ChIP. Cells are treated with formaldehyde to create DNA-protein-DNA cross-links. (Formaldehyde reacts with the amino groups on proteins and nucleic acids to form protein-protein and DNA protein covalent linkages). The DNA is then treated with a restriction nuclease that produces cohesive ends. Prior to the ligation step, the DNA is diluted so that the DNA ligase will join two different DNA fragments only if they are cross-linked. Finally, the cross-links are reversed and the DNA is purified, so that the ligated DNA molecules can be quantified by PCR. Are distant enhancers in proximity to the promoter? Chromosome conformation capture ( 3C) 3C reveals proximity of enhancers and promoters eg. colocalization in the ligated DNA product; eliminating transcription of a gene eliminates colocalization of enhancer and promoter sequences ( eg. b-globin locus)
1st example of 3C applied to enhancers: b globin locus:200kB A hypothetical model of the active chromatin hub (ACH) is shown to illustrate the 3D nature of the ACH (not to scale), not the actual position of the elements relative to each other in vivo. Red indicates the active regions (hypersensitive sites and active genes) of the locus forming a hub of hyperaccessible chromatin (ACH). The Inactive regions of the locus, having a more compact chromatin structure are indicated in gray, with the inactive h1 and y genes in lighter gray. Molecular Cell, Vol. 10, 1453–1465, December, 2002
5C applied to ES and NPC cells around key developmentally regulated genes to identify topologically associated domains (TAD) Cell 153, 1281–1295, June 6, 2013
Locus specific proteomics Identify proteins ( mass spectrometry) test their functional significance