Download

1 / 68
730 likes | 942 Views
The Anti-Tumor Function and Signal Transduction Pathway of p53 Protein. Introduction.

E N D
The Anti-Tumor Function and Signal Transduction Pathway of p53 Protein
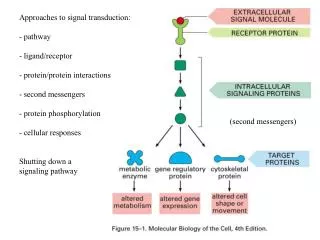
Introduction • P53 regulates>160 genes, acting as a tumor suppressor and maintaininggenome stability . Activated p53 either arrests the cellcycle to allow repair of damaged DNA or eliminates damaged cellsthrough apoptosis .
p53 has two modes of action for carrying out tumor suppression: • cell cycle arrest • apoptosis . • As previously mentioned, cell cycle arrest gives the cell time to repair the damaged DNA before cell division (mitosis) . p53 induced apoptosis occurs to protect against cancer development by eliminating potential tumorigenic cells .
Domain Arrangement • The structure ofp53 contains several domains: the acidic, transactivation, SH3-binding,core-DNA-binding, tetramerization, and regulatory domains. DNAbinding is critical for the biological functions of p53. Properp53–DNA binding requires a well folded core-binding domain(CBD) and a p53 homotetramer.
The N-terminal transactivation domain (N-term) contains 4 phosphorylation sites, Serine 15 is phosphorylated by ATM kinase and serine 20 can be phosphorylated by the cell check point kinase Chk2 . • The DNA binding domain (DBD) contains no posttranslational modifications. Any modification in this region would alter sequence-specific DNA binding. • The C-terminal domain is composed of a nuclear localization signal, a tetramerization domain, and a regulatory domain. Acetylation takes place in the C-terminal domain at 3 specific locations: Lysine 320 is acetylated by P/CAF, a histone acetyltransferase, while lysines 320 and 382 are acetylated by p300/CBP.
Modification of p53 • In response to cellular stress (DNA damage), the N-terminal activation domain of p53 is phosphorylated at serine 15 by ATM protein kinase, serines 20 by check point kinase chk2, phosphorylation of serine 20 is especially important in blocking HDM2 binding. • When p53 is bound to HDM2, it is not able to interact with TAFs (TATA Binding Protein Associated Factors) and no transcriptional activation will occur .
Phosphorylation of serine 15 has no effect on HDM2 binding, but does seem to increase the recruitment of p300 and CBP (CREB binding protein). • p300/CBP acetylate the C-terminal domain at lysine 373 and lysine 382, while lysine 320 is acetylated by the histone acetyltransferase P/CAF. • Acetylation is not required for p53 to bind to its promoter, but it may be important in stabilizing the protein and preventing cytoplasmic sequestering.
p53 induces cell cycle arrest • p53 induces cell cycle arrest at G1 by activating the transcription of p21. The cell cycle is stopped at G2 by both p21 and two other p53 transcriptional targets, 14-3-3sand Gadd45 (growth arrest inducible gene). • The three proteins inhibit the cyclin dependent kinase cdc2, which is essential for entry into mitosis. 14-3-3s anchors cdc2 in the cytoplasm, Gadd45 prevents the association of cdc2 with cyclin B1, and p21 inhibits cdc2 directly. • This method of cell cycle delay allows time for repair of DNA damage.
p53-dependent apoptosis • p53-dependent apoptosis is not as well understood yet. p53 is thought to promote apoptotic signaling in the cell by activating transcription of apopotosis-related genes ,such as BAX. • p53 is also able to promote apoptosis through transcription-independentapoptotic mechanisms. Under certain conditions, p53 inducesapoptosis in the absence of transcription or protein synthesis.
Extrinsic and intrinsic apoptotic pathways • The extrinsicpathway involves engagement of particular `death' receptorsthat belong to the tumor necrosis factor receptor (TNF-R) familyand, through the formation of the death-inducing-signaling-complex(DISC), leads to a cascade of activationof caspases, including caspase-8 and caspase-3, which in turninduce apoptosis.
The intrinsic pathway is triggered in responseto DNA damage and is associated with mitochondrial depolarizationand release of cytochrome c from the mitochondrial intermembranespace into the cytoplasm. • Cytochrome c, apoptotic protease-activatingfactor 1 (APAF-1) and pro-caspase-9 then form a complex termedthe apoptosome, in which caspase9 is activated and promotesactivation of caspase-3, caspase-6 and caspase-7.
Detailed extrinsic pathway • p53 can activate the extrinsic apoptotic pathway through theinduction of genes encoding three transmembranes proteins: • Fas,DR5 and PERP. • The cell-surface receptor Fas (CD95/Apo-1), amember of the TNF-R family of receptors, is a key componentof the extrinsic death pathway.
Fas is activated by binding of its ligand, FasL. p53 induces Fas mRNAexpression by binding to elements found in the promoter andfirst intron of the Fas gene. • In addition to stimulating Fas transcription, overexpressedp53 may enhance levels of Fas at the cell surface by promotingtrafficking of the Fas receptor from the Golgi
The second member of this receptor family that is induced byp53 is DR5/KILLER, the death-domain-containing receptor forTNF-related apoptosis-inducing ligand (TRAIL). • DR5 is inducedby p53 in response to DNA damage and in turnpromotes cell death through caspase-8.
Binding of TNF to TNFR1 results in receptor trimerisation and clustering of intracellular death domains. This allows binding of an intracellular adapter molecule called TRADD (TNFR-associated death domain) via interactions between death domains. TRADD has the ability to recruit a number of different proteins to the activated receptor. Recruitment of RIP (receptor interacting protein) and TRAF2 (TNF-associated factor 2) leads to activation of NF-kB and the JNK/Ap-1 pathway. • TRADD can also associate with FADD, which leads to the induction of apoptosis via the recruitment and cleavage of pro-caspase 8. • TNFR1 is also able to mediate apoptosis through the recruitment of an adapter molecule called RAIDD (RIP-associated ICH-1 / CED-3 homologous protein with a death domain). RAIDD associates with RIP through interactions between death domains and can recruit caspase 2 through an interaction with a motif, similar to the death effector domain, known as CARD (caspase recruitment domain). Recruitment of caspase 2 leads to induction of apoptosis.
The kineticsof PERP induction in response to DNA damage and the presenceof a p53-responsive element in the PERP promoter support thenotion that it is a direct p53 target. • A role for PERP in apoptosisis suggested by the significantly higher levels of PERP mRNAin cells undergoing apoptosis than in arresting cells.
The intrinsic pathway • The intrinsic apoptotic pathway is dominated by the Bcl-2 familyof proteins, • Intriguingly, a keysubset of the Bcl-2 family genes are p53 targets, includingBax, Noxa, PUMA and the most recently identified,Bid.
Bax was the first member of this group shown to be induced byp53, it resulted cytochromec releasesing from the mitochondria and caspase-9 activation . • Therequirement for Bax in p53-mediated apoptosis appears to becell-type dependent.
The PUMA gene is also directly induced by p53in response to DNA damage, through p53-responsive elements withinthe first intron of PUMA. • A vital balance between PUMA and p21 has beenidentified to determine the onset of arrest, or death, in responseto exogenous p53 expression .
PUMA expression promotes mitochondrialtranslocation and mulitmerization of Bax, culminating in apoptosisinduction • Bax participatesin the death response as an indirect target of p53 through PUMA
Noxaencodes a BH3-only proteinand hence is likely to contribute to p53-mediated apoptosisin a similar manner to PUMA and Bax, • Thus, it appears that, in response to DNA damage,p53 activates the intrinsic mitochondrial apoptotic pathwayby inducing the expression of at least three Bcl-2 pro-apoptoticfamily members, shifting the balance towards pro-apoptotic effects.
BID: a link between the extrinsic and intrinsic apoptotic pathways • The pro-apoptotic Bid is the only one found to link two pathway induced by p53. • Bid gene is transcriptionallyregulated by p53 through responseelements in the first intron of the human gene or in the promoterof the mouse gene.
Activation of Bid involves cleavageof cytoplasmic Bid by caspase-8 to expose a new N-terminal glycineresidue, which undergoes post-translational myristoylation. • Myristoylated Bid translocates to the mitochondria, insertsinto the membrane and activates BAX and BAK to initiate mitochondrialevents leading to apoptosome formation. • Some anti-tumor drugs were appears to be critically dependenton the presence of wild-type p53 and Bid.
PTEN-p53 • PTEN has also recentlybeen shown to physically interact with P53 , This interactionincreases the transcriptional activity of P53 and blocks itsdegradation. • Most of PTEN's major functions appear to be mediated by its lipid phosphatase activity, which antagonizes the effects of Class I PI3-kinases.
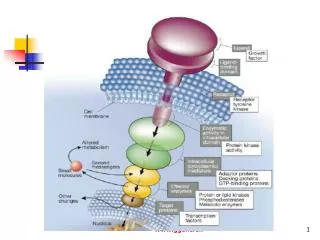
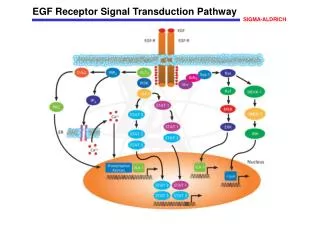
p53-mediated abrogation of survival signals: the AKT pathway • Binding of mitogens and cytokines to cell surface receptorsincluding the insulin receptor, the epidermal growth factorreceptor (EGFR) and the platelet-derived growth factor receptor(PDGFR), and the actions of oncogenes such as Ras and Her2/Neu,is transduced by phosphoinositide 3-kinase (PI3K) activatingsignaling pathways that promote cell proliferation and viability
PI3K activates AKT,a serine/threonine kinase, through phosphorylation on Ser473.In turn, AKT phosphorylates a range of targetsthat function to promote cell survival, including the majorinhibitor of p53, Mdm2. • This phosphorylation enhances the nuclear accumulation of Mdm2,augments Mdm2 interaction with p300, and reduces the affinityof Mdm2 for p19ARF. • Consequently, AKT augments the inhibition and destabilizationof p53 by Mdm2.
stress-induced activationof p53 counteracts the inhibitory effects of this survival pathwayby multiple mechanisms • First, p53 promotes caspase-mediatedcleavage and subsequent degradation of the AKT protein itself. • Second, p53 induces the expressionof the PTEN tumor suppressor gene, which encodes a phosphatasethat dephosphorylates PI3K, thereby impairing AKT activation. • Third, p53 induces expressionof cyclin G, which in turn recruits the phosphatase PP2AB' tothe Mdm2-p53 complex, where it dephosphorylates Mdm2 at theAKT phosphorylation sites.
These feedback loops determine thesurvival versus apoptotic outcome in the interplay between p53and the AKT survival pathway. • This fine balance is often interrupted in cancer, eitherby mutations in PTEN or amplification of Mdm2.
Caspase Activity • p53boosts the activation of the caspase cascade by both transcription-dependentand -independent mechanisms. • Transcription -independent mechanisms. Caspase-8 can contribute to, although is not alwaysessential for, DNA-damaged induced death.
Transcription-dependent mechanisms • In response to DNA damage, p53 directly induces caspase-6 expressionthrough a response element within the third intron of the gene. Caspase-6 cleaves the nuclearenvelope protein lamin A and several transcription factors. • Caspase-6 plays an important role in p53-inducedneuronal cell death and is the major protein involved in thecleavage of the amyloid precursor protein.
How p53 determinating cell fate • Exacting discrimination between p53 arrest and apoptotic functionshas been critical to the identification of the importance ofthe latter in tumor suppression. A strong link between the apoptoticfunction of p53 and tumor suppression has been demonstrated
Several generalfactors that influence this decision include p53 expressionlevels, the type of stress signal, the cell type and the cellularcontext at the time of exposure to stress. • Present study supports the notion that levels of genotoxicagent introduced to cells may determine in a p53-dependent mannerwhether cells will undergo apoptosis or induce DNA repair
In response to low and moderate levels of DNA damage,p53 protein activates the DNA repair machinery represented byBER. This is probably by direct participationof p53 in the repair complex. • On the other hand, high levelsof DNA damage may induce different p53 post-translational modificationsthat down regulate BER pathway and in turn induce an apoptoticresponse.