Download

1 / 53
550 likes | 865 Views
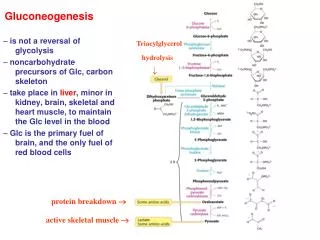
Gluconeogenesis. Some tissues, such as brain, RBCs, kidney medulla, testes, embrionic tissues and exercising muscle require a continuous supply of glucose for metabolic energy. The human brain requires over 120 gm of glucose per day. Mammalian cells make glucose from simpler precursors.

E N D
Gluconeogenesis • Some tissues, such as brain, RBCs, kidney medulla, testes, embrionic tissues and exercising muscle require a continuous supply of glucose for metabolic energy. • The human brain requires over 120 gm of glucose per day. • Mammalian cells make glucose from simpler precursors. • Liver glycogen can meet these needs for only 10 to 18 hours without dietary carbohydrate.
During a prolonged fast, • Hepatic liver stores are depleted, glc is formed from other molecules, such as • Lactate • Pyruvate • Glycerol • Alpha keto acids • The formation of glc from nonhexose precursors is called gluconeogenesis (formation of new sugar).
Pyruvate precursors • The direct Glc reserves are sufficient to meet Glc needs for about a day! • Gluconeogenic pathway makes Glc from pyruvate precursors. • Triacyl glycerol Glycerol + Fatty acids • Glycerol is a precursor of glc, glycerol enters glycolytic pathway as dihydroxyacetone phosphate.
Gluconeogenesis is NOT a reversal of glycolysis • Several reactions MUST differ because of the irreversible steps. • HK (hexokinase) • PFK (phosphofructokinase) • PK (pyruvatekinase)
Let’s make Glc from pyruvate • Carboxylation of pyruvate Pyruvate + CO2 + ATP + H2O OA +ADP + Pi + 2H • Enzyme: Pyruvatecarboxylase OA + GTP PEP+ GDP + CO2 • Enzyme: PEP-carboxykinase
Domain structure of pyruvatecarboxylase • ATP grasp: • Activates bicarbonate ions and transfers CO2 to the biotin domain. • From there, CO2 is transferred to pyruvate.
Carboxylation of pyruvate • Pyruvate carboxylase contains BIOTIN, which is covalently bound to the enzyme through lysine • Enzyme + CO2 + ATP-----> Carboxybiotin-enzyme +ADP +Pi • Carboxybiotin-enzyme + pyruvate------->OA + Enzyme • BIOTIN carries CO2...
Biotin is covalently attached group • Biotin serves as a carrier for activated CO2. • e-amino group and carboxylate group of biotin are linked. • CO2 is found mainly as HCO3 in our system. • When Acetyl CoA is high, then biotin is carboxylated. • The activated carboxyl group is transferred from carboxybiotin to pyruvate to form oxaloacetate.
2. Transport of OA to the cytoplasm • Pyuvatecarboxylase is a mitochondrial enzyme, whereas the other enzymes in gluconeogenesis are cytoplasmic. • OA should be transported to the cytoplasm. • How? • It is reduced to MALATE first and then transferred to the cytoplasm. • In the cytoplasm, it is reoxidized to OA.
3. Decarboxylation of cytoplasmic OA • OA is decarboxylated and P-lated by PEP carboxykinase in the cytosol (PEP is made then!) • The overall reaction catalyzed by the combined action of pyruvatecarboxylase and PEP carboxykinase provides a pathway from Pyruvate PEP. • Therefore, once PEP is formed, it enters the reversed reactions of glycolysis until it reaches F-1,6 Bisphosphate!
4. Dephosphorylation of F-1,6BP Fructose 1,6-bisphosphate + H2O F-6-P + Pi • Enzyme: Fructose1,6-bisphosphatase • This enzyme plays an important role in regulation. • It is inhibited by F 2,6 BisP, an allosteric modifier whose concentration is influenced by the levels of circulating glucagon. • This enzyme is found in liver and kidney.
5. Generation of free Glc Dephospharylation of Glc 6-P Glc 6-P + H2O D-Glc + Pi • Enzyme: Glc 6-phosphatase • It is found in liver and kidney but not in muscle and brain. • Thus, muscle and brain cannot make Glc by gluconeogenesis • Type I glycogen storage disease results from an inherited deficiency of glc 6-phosphatase.
Freeing Glc • The final step, freeing Glc, takes place in ER lumen where it is hydrolyzed to Glc by Glc 6-Phosphatase, a membrane bound enzyme. • Calcium binding protein (SP) is necessary for phosphatase activity. • Glc and Pi are shuttled back to the cytosol by a pair of transporters. • The glucose transporter in the ER membrane is like those found in the plasma membrane.
Gluconeogenesis is energetically costly! • The stoichiometry of gluconeogenesis is: 2 pruvate + 4 ATP + 2 GTP + 2 NADH + 6 H2O Glc + 4 ADP + 2 GDP + 6 Pi + 2 NAD+ + 2 H+ • In contrast, the stoichiometry of reversal of glycolysis is: 2 pyruvate + 2 ATP + NADH + 2 H2O Glc + 2 ADP + 2 Pi + 2 NAD+ + 2 H+ • The difference is 4 ATP. This is needed to turn energetically unfavorable process to a favorable one!
Gluconeogenesis and glycolysis are reciprocally regulated • Both glycolysis and gluconeogenesis are highly exorgonic under cellular conditions so there is no thermodynamic barrier. • But, amounts and activities of the distinctive enzymes of each pathway are controlled so that both pathways are not highly active at the same time.
Substrate cycles F-6-P F 1,6BisP • A pair of reactions such as the above one is called “substrate cycle” • There is also some cycling in irreversible reactions. • “Imperfection” in metabolism? • They are sometimes referred as “futile cycles” • Futile cycles amplify metabolic signals! • The other potential biological role of substrate cycles is the generation of heat produced by the hydrolysis of ATP.
Lactate and alanine formed by contracting muscle are used by other organs • Lactate is a dead end in metabolism. • Lactate should be converted to pyruvate. • The plasma membranes of most cells are highly permeable to lactate and pyruvate; therefore, they easily diffuse to go to liver! • Excess lactate enters the liver and is converted to pyruvate first and then to glucose. • Thus, the liver restores the level of glucose necessary for active muscle cells, which derive ATP from the glycolytic conversion of glucose into lactate. Contracting skeletal muscle supplies lactate to the liver, which uses it to make glucose. • These reactions constitute the CORI CYCLE.
LDH enzyme • Lactate Pyruvate by LDH (lactate dehydrogenase). • The interconversion of pyruvate and lactate are done by different subunits of LDH. LDH is a tetramer. • H in the heart • M in the muscle
REGULATION 1. Control point:Pyruvatecarboxylase • Acetyl CoA is a + allosteric modulator for the pyruvatecarboxylase enzyme. • Glc is made from pyruvate when there is a lot of Acetyl CoA (more Acetyl CoA than TCA cycle can handle) • Acetyl CoA inhibits the pyruvatedehydrogenase enzyme but stimulates the pyruvatecarboxylase. 2. Control point: F 1,6 bisphoshatase reaction 3.Control point: F-2,6 bisphosphate • Hormonal control
Hormonal Control • The special role of the liver is to maintain constant blood glucose level and requires additional control mechanisms. • When blood glucose decreases, glucagon increases and glucose is released. • This hormonal regulation in the liver is mediated by fructose-2,6-bisphosphate, which is a allostericeffector for PFK-1, and F-1,6-bisphosphate
Role of F2,6BP in regulation of Glycolysis and Gluconeogenesis Strongly stimulates PFK-1 Strongly inhibits FBPase-1